

The inflammatory effects of UDP-glucose in N9 microglia are not mediated by P2Y14 receptor activation
- PMID: 18368535
- PMCID: PMC2246002
- DOI: 10.1007/s11302-008-9095-1
The inflammatory effects of UDP-glucose in N9 microglia are not mediated by P2Y14 receptor activation
Abstract
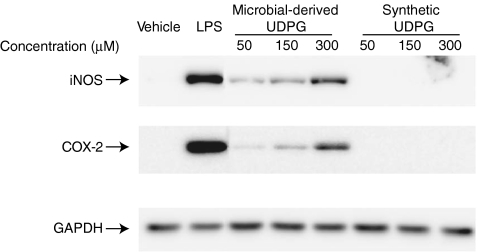
In this study we evaluated the functionality and inflammatory effects of P2Y14 receptors in murine N9 microglia. The selective P2Y14 receptor agonist UDP-glucose (UDPG) derived from microbial sources dose dependently stimulated expression of cyclooxygenase-2 and inducible nitric oxide synthase, and potentiated the effects of bacterial lipopolysaccharide on nitric oxide production. However, another selective P2Y14 receptor agonist, UDP-galactose, did not affect these endpoints either alone or in combination with lipopolysaccharide. Interestingly, synthetic UDPG also had no detectable pro-inflammatory effects, although P2Y14 receptors are both expressed and functional in N9 microglia. While synthetic UDPG decreased levels of phosphorylated cyclic AMP response element binding protein, an effect that was blocked by pertussis toxin, the pro-inflammatory effects of microbial-derived UDPG were insensitive to pertussis toxin. These data suggest that the pro-inflammatory effects of microbial-derived UDPG are independent of P2Y14 receptors and imply that microbial-derived contaminants in the UDPG preparation may be involved in the observed inflammatory effects.
Figures
References
-
- Abbracchio MP, Boeynaems J-M, Barnard EA, Boyer JL, Kennedy C, Miras-Portugal MT, King BF, Gachet C, Jacobson KA, Weisman GA, Burnstock G (2003) Characterization of the UDP-glucose receptor (re-named here the P2Y14 receptor) adds diversity to the P2Y receptor family. Trends Pharmacol Sci 24(2):52 - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
