

Identification of critical residues in Gap3 of Streptococcus parasanguinis involved in Fap1 glycosylation, fimbrial formation and in vitro adhesion
- PMID: 18371226
- PMCID: PMC2335109
- DOI: 10.1186/1471-2180-8-52
Identification of critical residues in Gap3 of Streptococcus parasanguinis involved in Fap1 glycosylation, fimbrial formation and in vitro adhesion
Abstract
Background: Streptococcus parasanguinis is a primary colonizer of human tooth surfaces and plays an important role in dental plaque formation. Bacterial adhesion and biofilm formation are mediated by long peritrichous fimbriae that are composed of a 200 kDa serine rich glycoprotein named Fap1 (fimbriae-associated protein). Glycosylation and biogenesis of Fap1 are modulated by a gene cluster downstream of the fap1 locus. A gene encoding a glycosylation-associated protein, Gap3, was found to be important for Fap1 glycosylation, long fimbrial formation and Fap1-mediated biofilm formation.
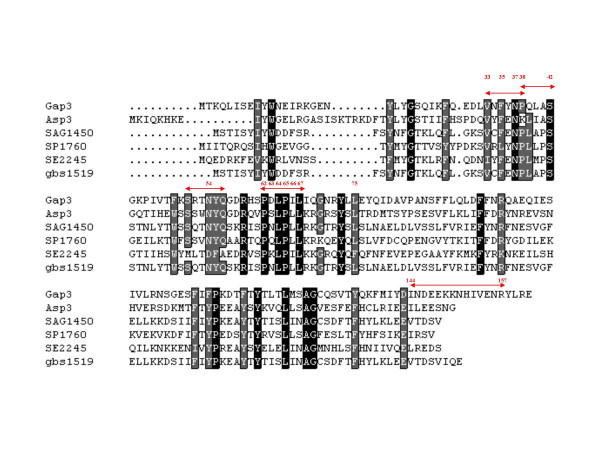
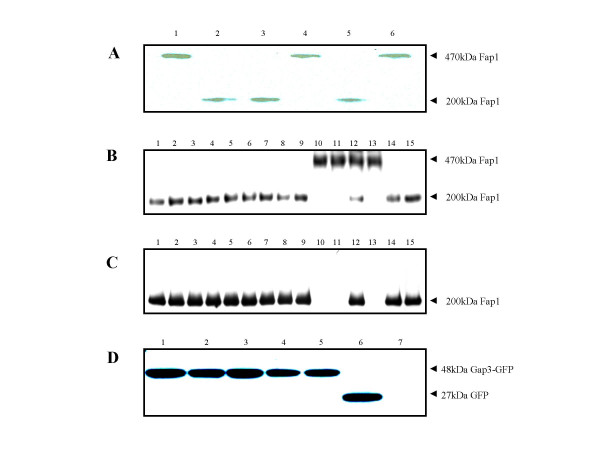
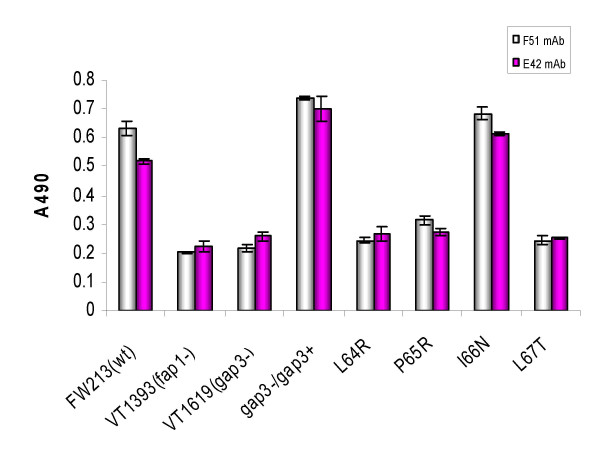
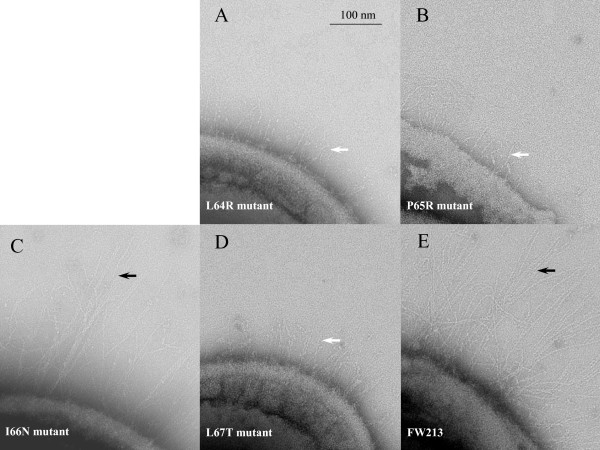
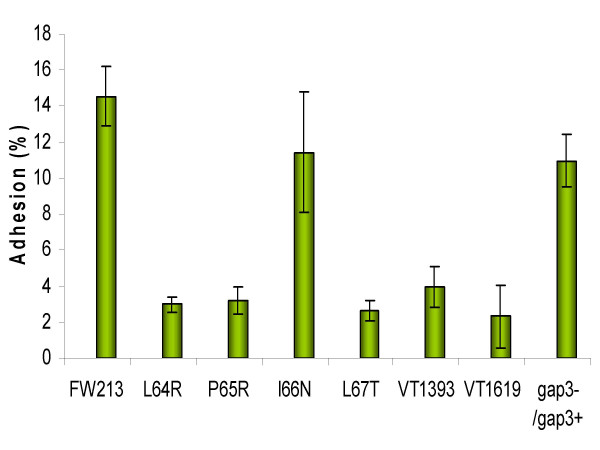
Results: Deletion and site-directed mutagenesis were employed to dissect the regions within Gap3 that were important for its function in Fap1 glycosylation and biogenesis. A deletion of 6 consecutive amino acids, PDLPIL, eliminated the production of the mature 200 kDa Fap1 protein and gave rise instead to a 470 kDa Fap1 intermediate that was only partially glycosylated. Site-directed mutagenesis of the 6 amino acids revealed that only three of these amino acids were required. Mutants in these amino acids (L64R, P65R and L67T) produced the premature 470 kDa Fap1 intermediate. Mutants in the remaining amino acids produced the mature form of Fap1. Cell surface expression of the Fap1 precursor among L64R, P65R and L67T mutants was reduced to levels consistent with that of a gap3 insertional mutant. Electron micrographs showed that these 3 mutants lost their long peritrichous fimbriae. Furthermore, their in vitro adhesion ability to saliva-coated hydroxylapatite (SHA) was inhibited.
Conclusion: Our data suggest that 3 highly conserved, hydrophobic residues L64, P65 and L67 in Gap3 are essential for Gap3 function and are important for complete glycosylation of Fap1, fimbrial formation and bacterial adhesion.
Figures
Similar articles
-
Role of gap3 in Fap1 glycosylation, stability, in vitro adhesion, and fimbrial and biofilm formation of Streptococcus parasanguinis.Oral Microbiol Immunol. 2008 Feb;23(1):70-8. doi: 10.1111/j.1399-302X.2007.00401.x. Oral Microbiol Immunol. 2008. PMID: 18173801
-
A conserved C-terminal 13-amino-acid motif of Gap1 is required for Gap1 function and necessary for the biogenesis of a serine-rich glycoprotein of Streptococcus parasanguinis.Infect Immun. 2008 Dec;76(12):5624-31. doi: 10.1128/IAI.00534-08. Epub 2008 Oct 13. Infect Immun. 2008. PMID: 18852249 Free PMC article.
-
A conserved domain of previously unknown function in Gap1 mediates protein-protein interaction and is required for biogenesis of a serine-rich streptococcal adhesin.Mol Microbiol. 2008 Dec;70(5):1094-104. doi: 10.1111/j.1365-2958.2008.06456.x. Epub 2008 Sep 30. Mol Microbiol. 2008. PMID: 18826412 Free PMC article.
-
Molecular strategies for fimbrial expression and assembly.Crit Rev Oral Biol Med. 2001;12(2):101-15. doi: 10.1177/10454411010120020101. Crit Rev Oral Biol Med. 2001. PMID: 11345521 Review.
-
Assembly of pili in Gram-positive bacteria.Trends Microbiol. 2004 May;12(5):228-34. doi: 10.1016/j.tim.2004.03.004. Trends Microbiol. 2004. PMID: 15120142 Review. No abstract available.
Cited by
-
The sweet tooth of bacteria: common themes in bacterial glycoconjugates.Microbiol Mol Biol Rev. 2014 Sep;78(3):372-417. doi: 10.1128/MMBR.00007-14. Microbiol Mol Biol Rev. 2014. PMID: 25184559 Free PMC article. Review.
-
Exome capture from saliva produces high quality genomic and metagenomic data.BMC Genomics. 2014 Apr 4;15:262. doi: 10.1186/1471-2164-15-262. BMC Genomics. 2014. PMID: 24708091 Free PMC article.
-
Sulfoquinovose synthase - an important enzyme in the N-glycosylation pathway of Sulfolobus acidocaldarius.Mol Microbiol. 2011 Dec;82(5):1150-63. doi: 10.1111/j.1365-2958.2011.07875.x. Epub 2011 Nov 7. Mol Microbiol. 2011. PMID: 22059775 Free PMC article.
-
A role for glycosylated serine-rich repeat proteins in gram-positive bacterial pathogenesis.Mol Oral Microbiol. 2012 Aug;27(4):257-69. doi: 10.1111/j.2041-1014.2012.00653.x. Epub 2012 Jun 11. Mol Oral Microbiol. 2012. PMID: 22759311 Free PMC article. Review.
-
Similarities and differences in the glycosylation mechanisms in prokaryotes and eukaryotes.Int J Microbiol. 2010;2010:148178. doi: 10.1155/2010/148178. Epub 2011 Jan 27. Int J Microbiol. 2010. PMID: 21490701 Free PMC article.
References
-
- Cisar JO, Brennan MJ, Sandberg AL. Lectin-specific Interaction of Actinomyces Fimbriae with Oral Streptococci. Washington, DC: American Society for Microbiology Press; 1985.
-
- Jenkinson HF, Lamont RJ. Streptococcal adhesion and colonization. Crit Rev Oral Biol Med. 1997;8:175–200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
