

The p47 GTPase Lrg-47 (Irgm1) links host defense and hematopoietic stem cell proliferation
- PMID: 18371424
- PMCID: PMC2278017
- DOI: 10.1016/j.stem.2007.10.007
The p47 GTPase Lrg-47 (Irgm1) links host defense and hematopoietic stem cell proliferation
Abstract
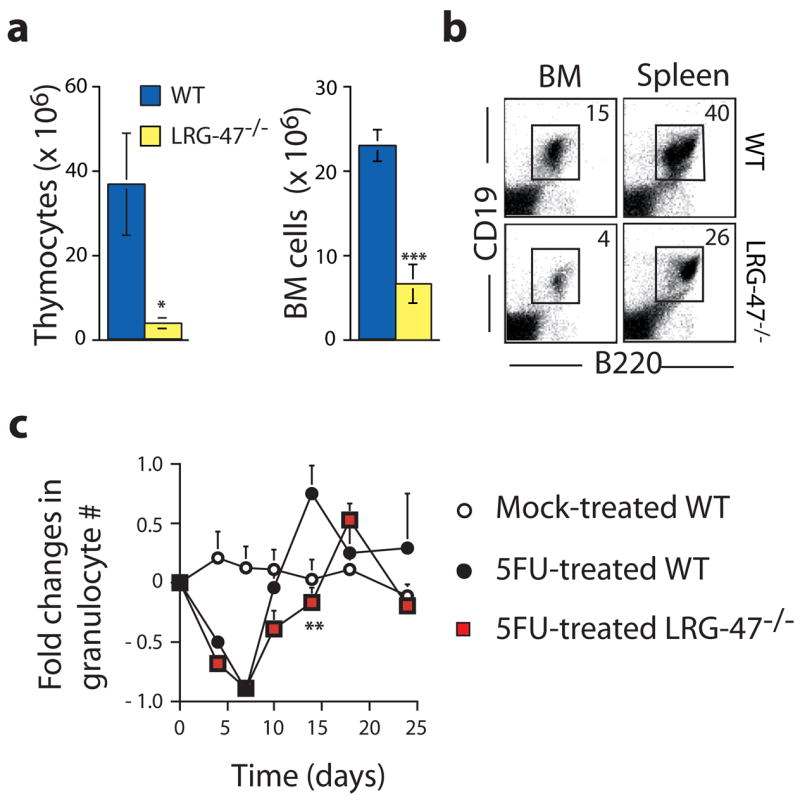
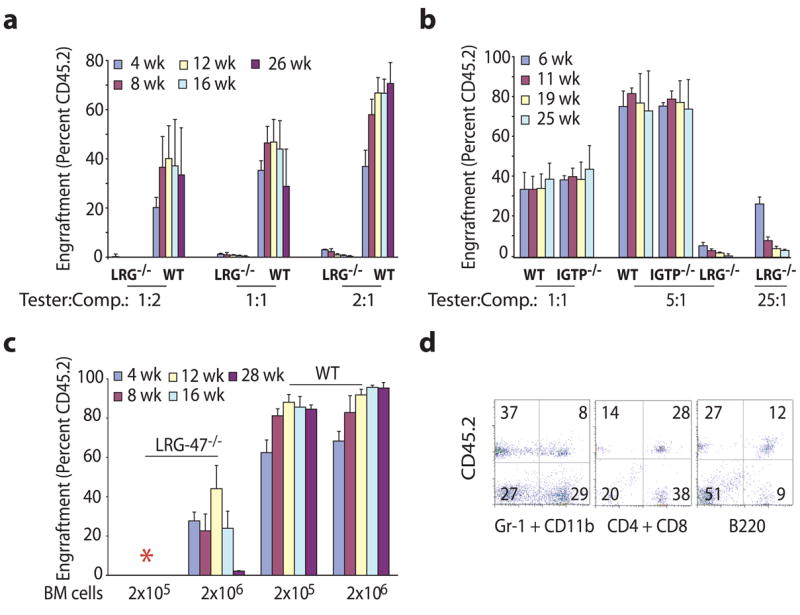
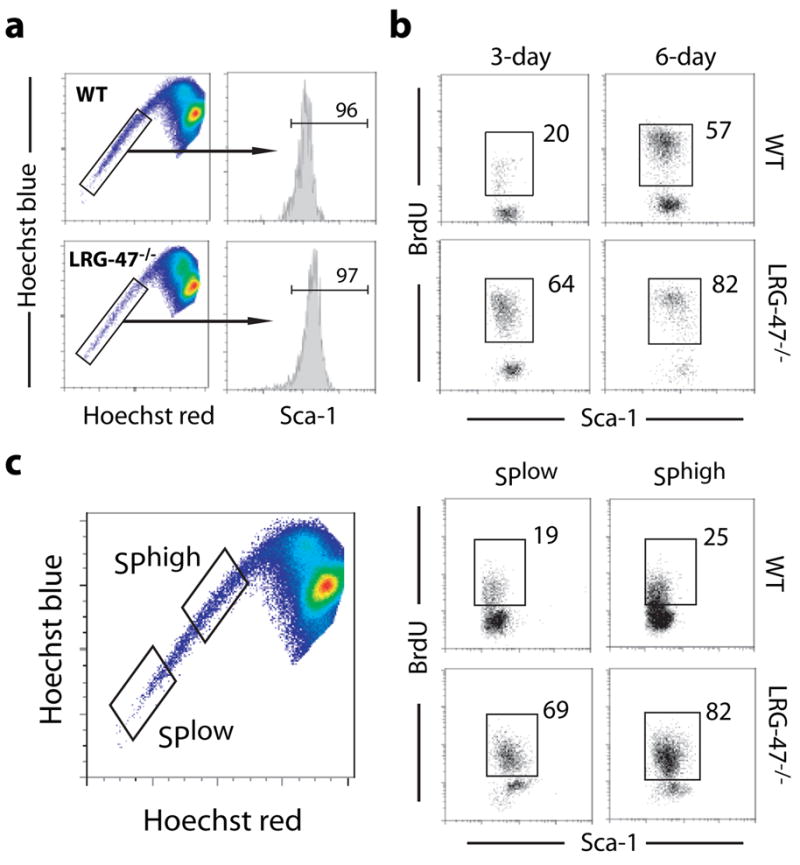
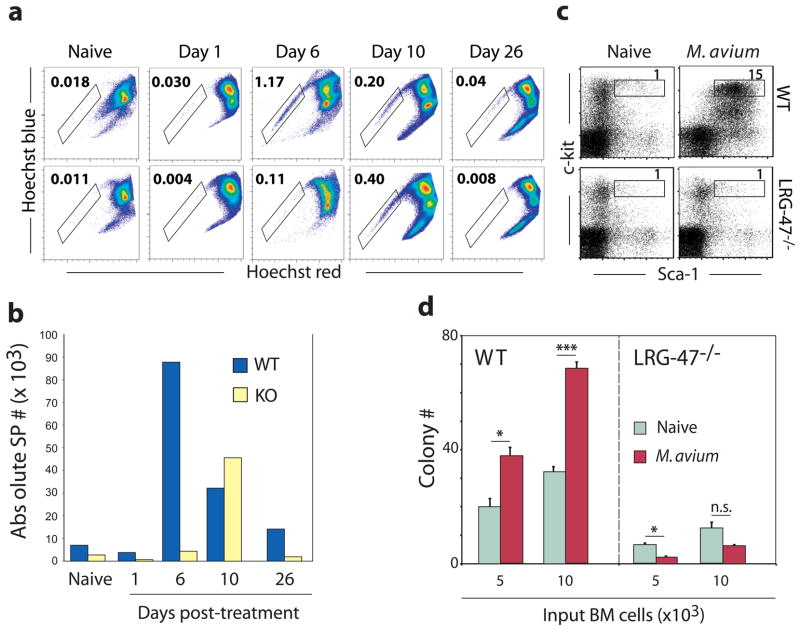
Hematopoietic stem cells (HSCs) are self-renewing bone marrow cells that give rise to all blood lineages and retain a remarkable capacity to proliferate in response to insult. Although some controls on HSC activation are known, little is understood about how this process is linked to natural signals. We report that the interferon-inducible GTPase Lrg-47 (Irgm1), previously shown to play a critical role in host defense, inhibits baseline HSC proliferation and is required for a normal HSC response to chemical and infectious stimuli. Overproliferating Lrg-47(-/-) HSCs are severely impaired in functional repopulation assays, and when challenged with hematopoietic ablation by 5-fluorouracil or infection with Mycobacterium avium, Lrg-47(-/-) mice fail to achieve the expected expansion response in stem and progenitor cell populations. Our results establish a link between the response to infection and HSC activation and demonstrate a novel function for a member of the p47 GTPase family.
Figures
Comment in
-
Stem cell regulation and host defense: the logic and the paradox.Cell Stem Cell. 2008 Jan 10;2(1):1-2. doi: 10.1016/j.stem.2007.12.002. Cell Stem Cell. 2008. PMID: 18371410
References
-
- Dufour C, Corcione A, Svahn J, Haupt R, Battilana N, Pistoia V. Interferon gamma and tumour necrosis factor alpha are overexpressed in bone marrow T lymphocytes from paediatric patients with aplastic anaemia. Br J Haematol. 2001;115:1023–1031. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
