

Lipidomic analysis and electron transport chain activities in C57BL/6J mouse brain mitochondria
- PMID: 18373617
- PMCID: PMC3104050
- DOI: 10.1111/j.1471-4159.2008.05383.x
Lipidomic analysis and electron transport chain activities in C57BL/6J mouse brain mitochondria
Abstract
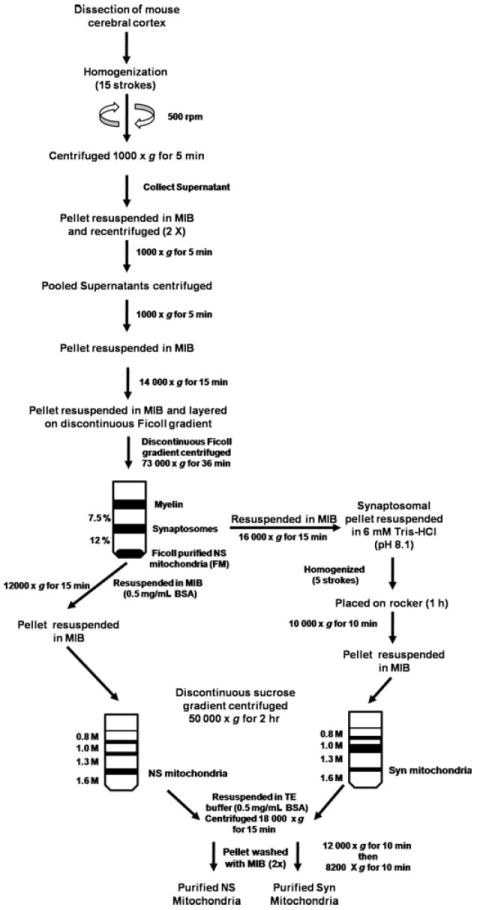
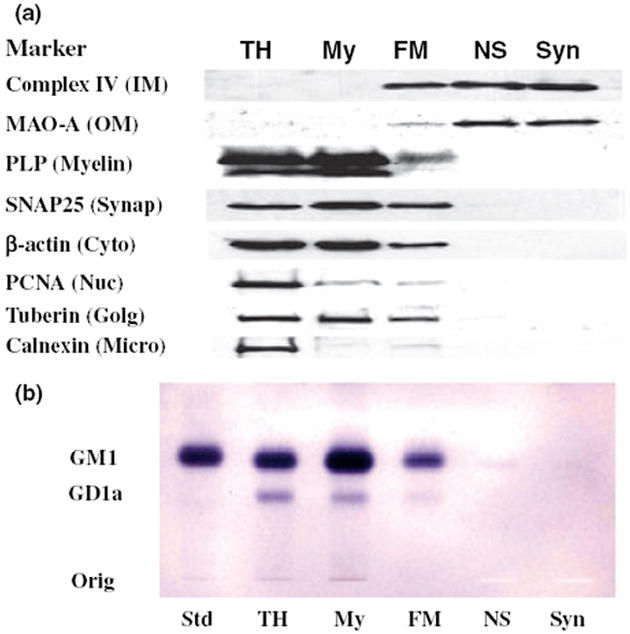
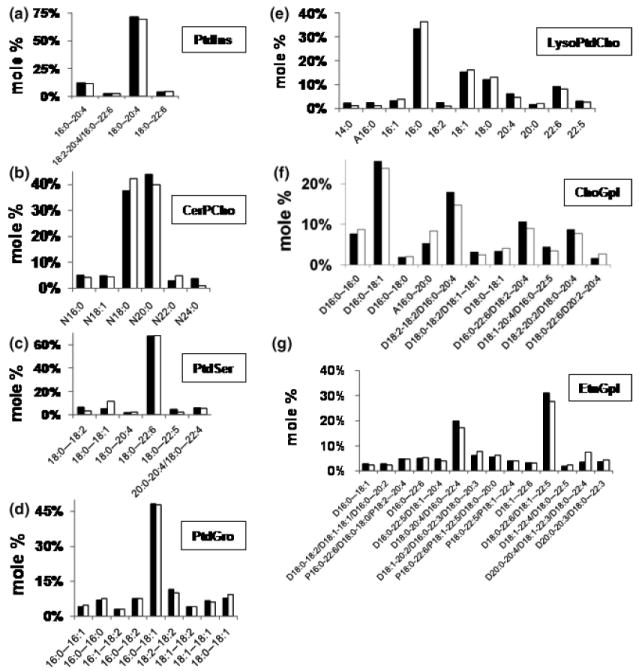
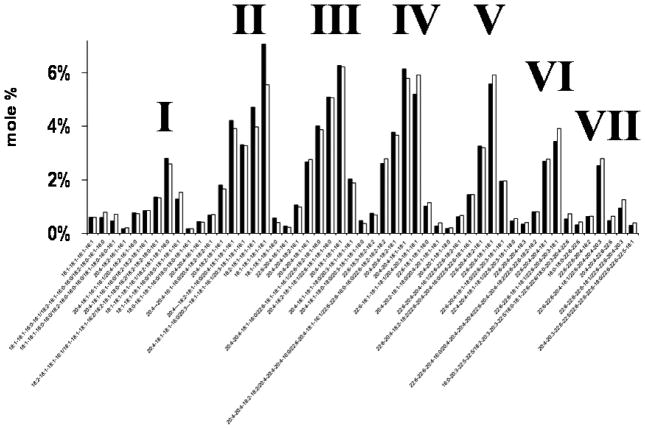
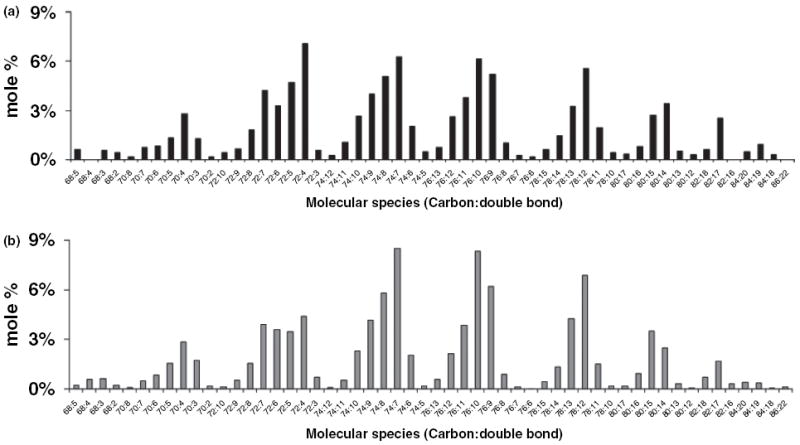
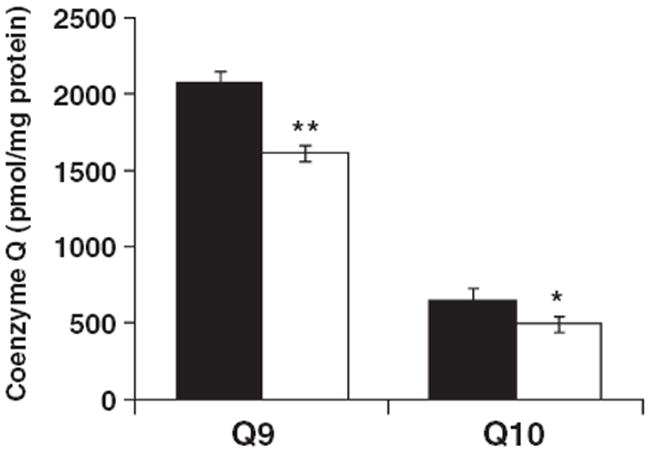
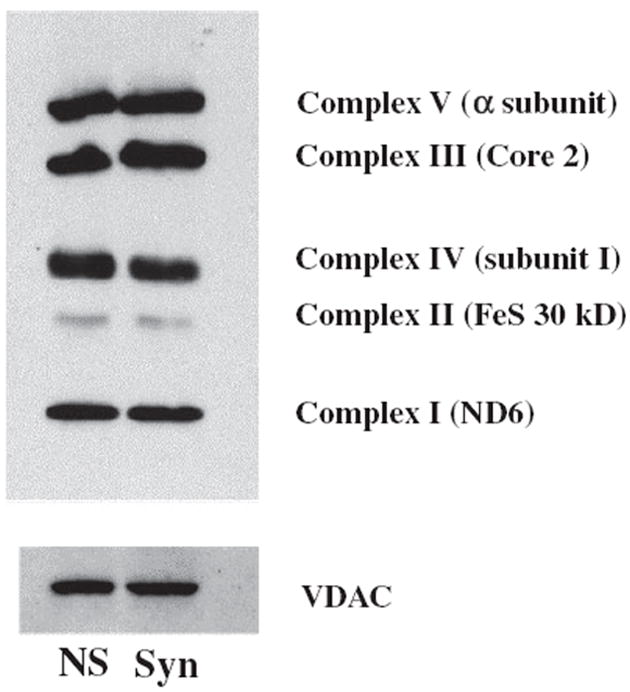
The objective of this study was to characterize the lipidome and electron transport chain activities in purified non-synaptic (NS) and synaptic (Syn) mitochondria from C57BL/6J mouse cerebral cortex. Contamination from subcellular membranes, especially myelin, has hindered past attempts to accurately characterize the lipid composition of brain mitochondria. An improved Ficoll and sucrose discontinuous gradient method was employed that yielded highly enriched mitochondrial populations free of myelin contamination. The activities of Complexes I, II, III, and II/III were lower in Syn than in NS mitochondria, while Complexes I/III and IV activities were similar in both populations. Shotgun lipidomics showed that levels of cardiolipin (Ptd(2)Gro) were lower, whereas levels of ceramide and phosphatidylserine were higher in Syn than in NS mitochondria. Coenzyme Q(9) and Q(10) was also lower in Syn than in NS mitochondria. Gangliosides, phosphatidic acid, sulfatides, and cerebrosides were undetectable in brain mitochondria. The distribution of Ptd(2)Gro molecular species was similar in both populations and formed a unique pattern, consisting of seven major molecular species groups, when arranged according to mass to charge ratios. Remodeling involving choline and ethanolamine phosphoglycerides could explain Ptd(2)Gro heterogeneity. NS and Syn mitochondrial lipidomic heterogeneity could influence energy metabolism, which may contribute to metabolic compartmentation of the brain.
Figures
References
-
- Battino M, Bertoli E, Formiggini G, Sassi S, Gorini A, Villa RF, Lenaz G. Structural and functional aspects of the respiratory chain of synaptic and nonsynaptic mitochondria derived from selected brain regions. J Bioenerg Biomembr. 1991;23:345–363. - PubMed
-
- Battino M, Gorini A, Villa RF, Genova ML, Bovina C, Sassi S, Littarru GP, Lenaz G. Coenzyme Q content in synaptic and non-synaptic mitochondria from different brain regions in the ageing rat. Mech Ageing Dev. 1995;78:173–187. - PubMed
-
- Bayir H, Tyurin VA, Tyurina YY, et al. Selective early cardiolipin peroxidation after traumatic brain injury: an oxidative lipidomics analysis. Ann Neurol. 2007;62:154–169. - PubMed
-
- Bedell MA, Jenkins NA, Copeland NG. Mouse models of human disease Part I: techniques and resources for genetic analysis in mice. Genes Dev. 1997a;11:1–10. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
