

Formation of the factory matrix is an important, though not a sufficient function of nonstructural protein mu NS during reovirus infection
- PMID: 18374384
- PMCID: PMC2486453
- DOI: 10.1016/j.virol.2008.02.024
Formation of the factory matrix is an important, though not a sufficient function of nonstructural protein mu NS during reovirus infection
Abstract
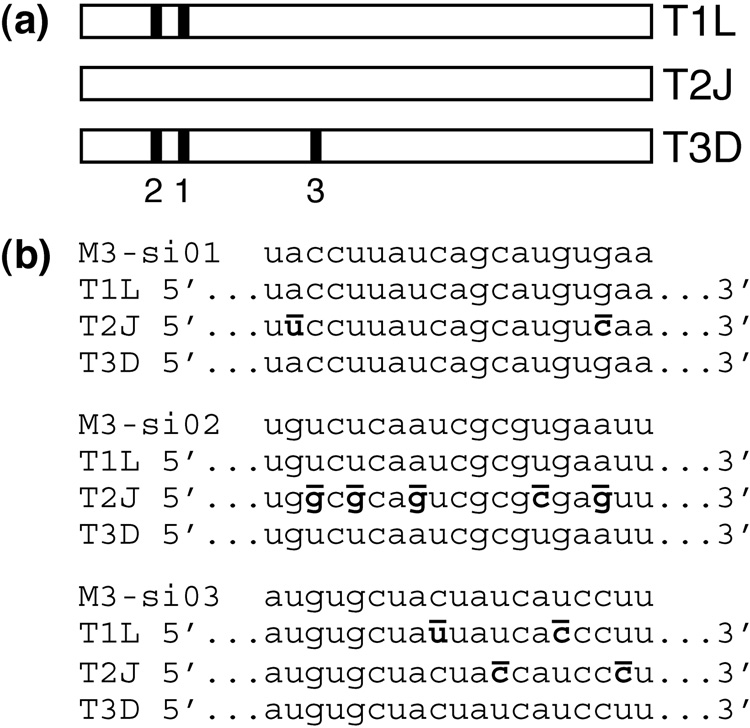
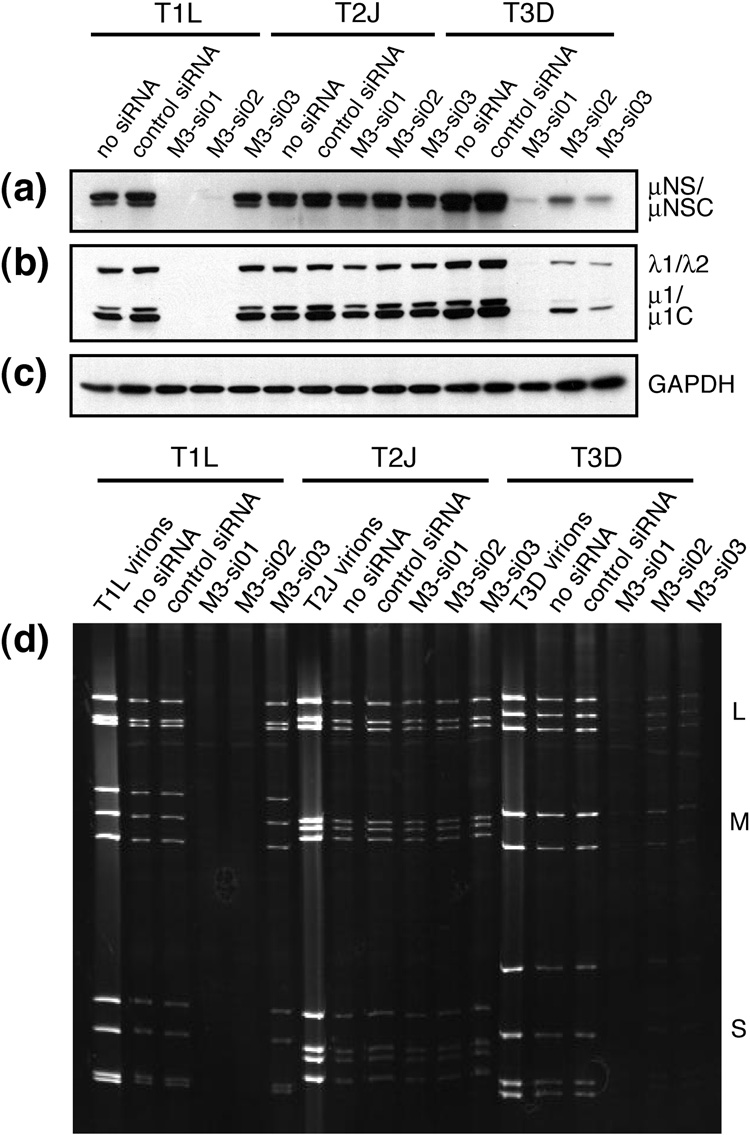
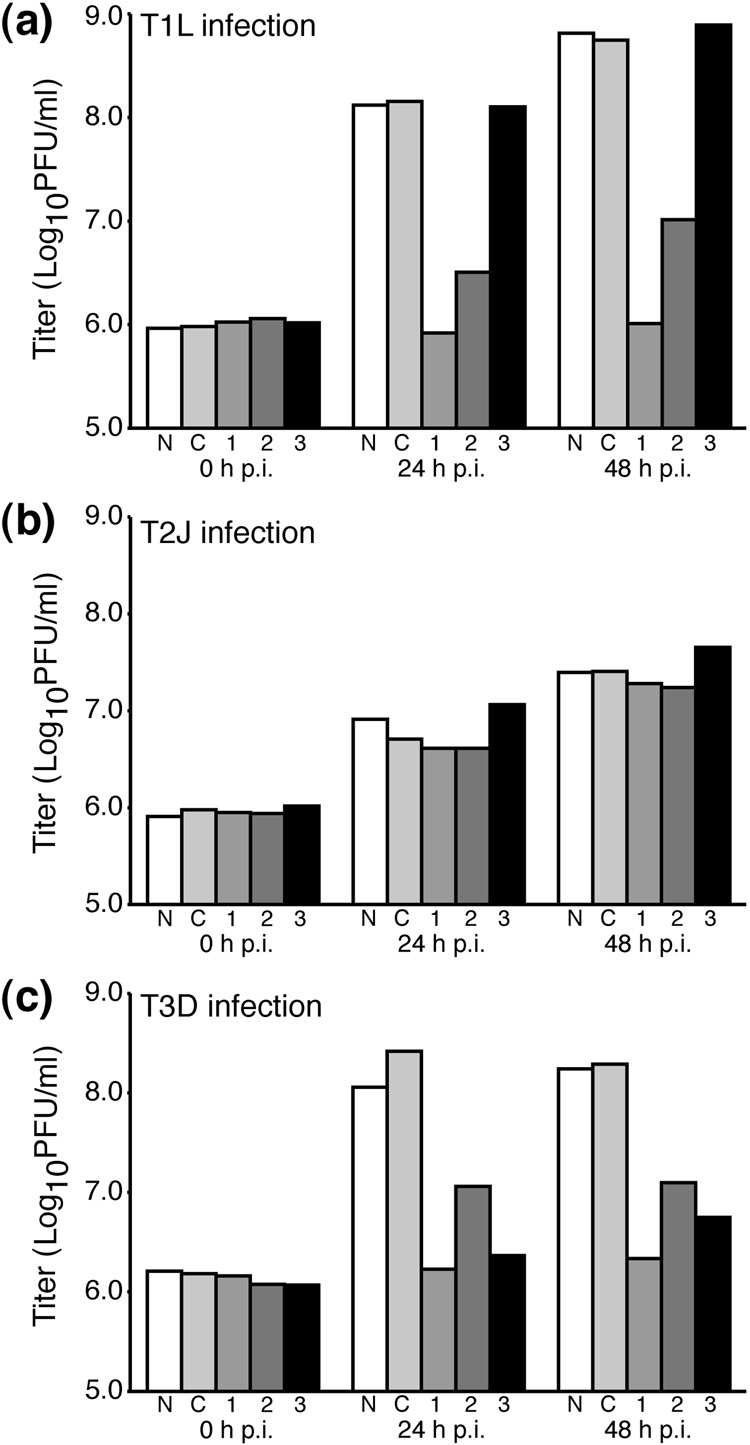
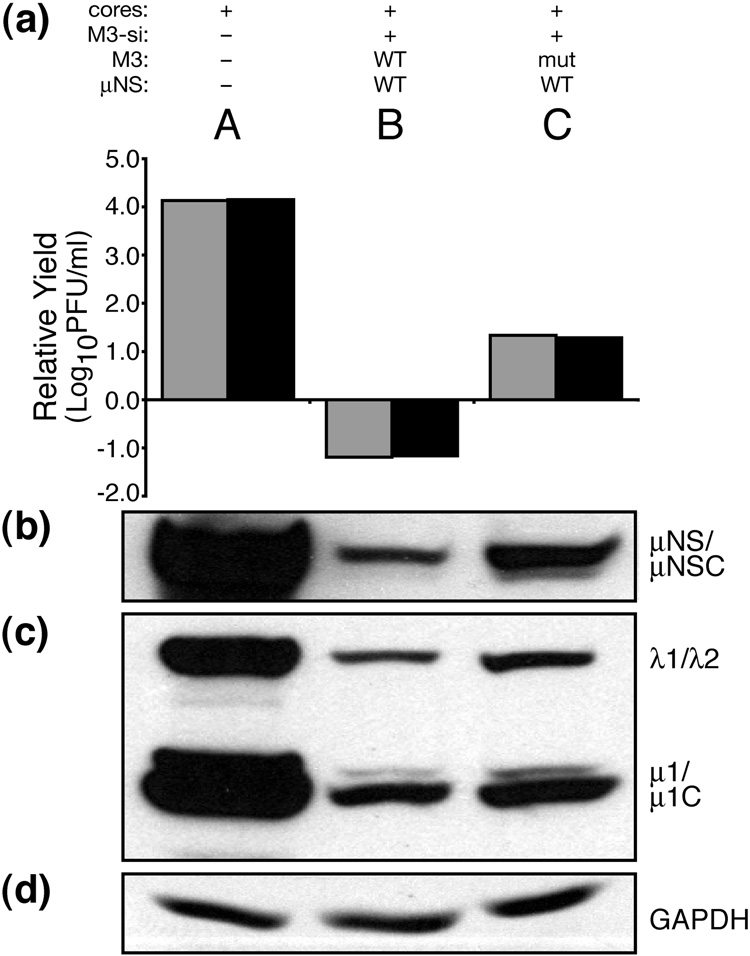
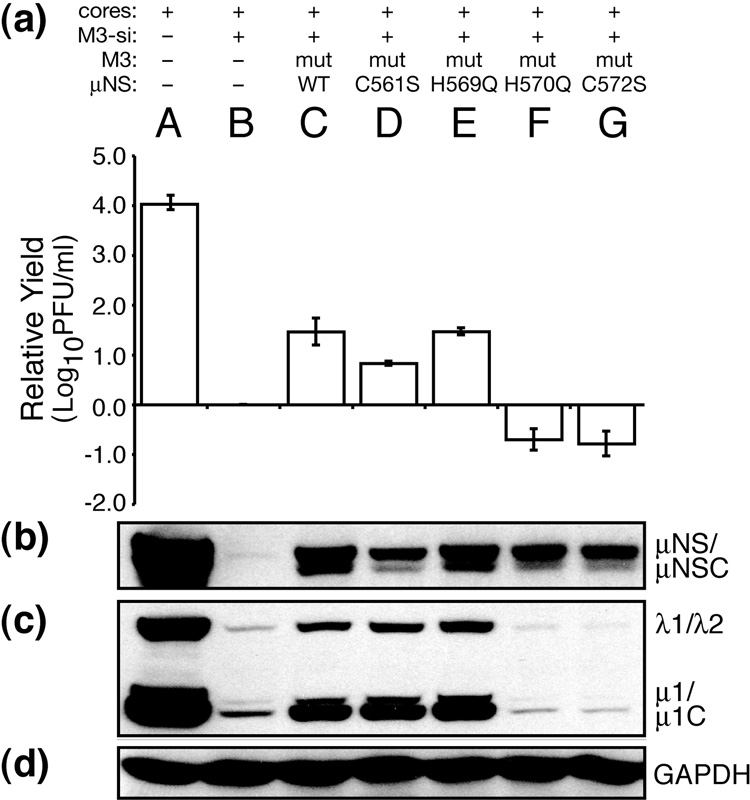
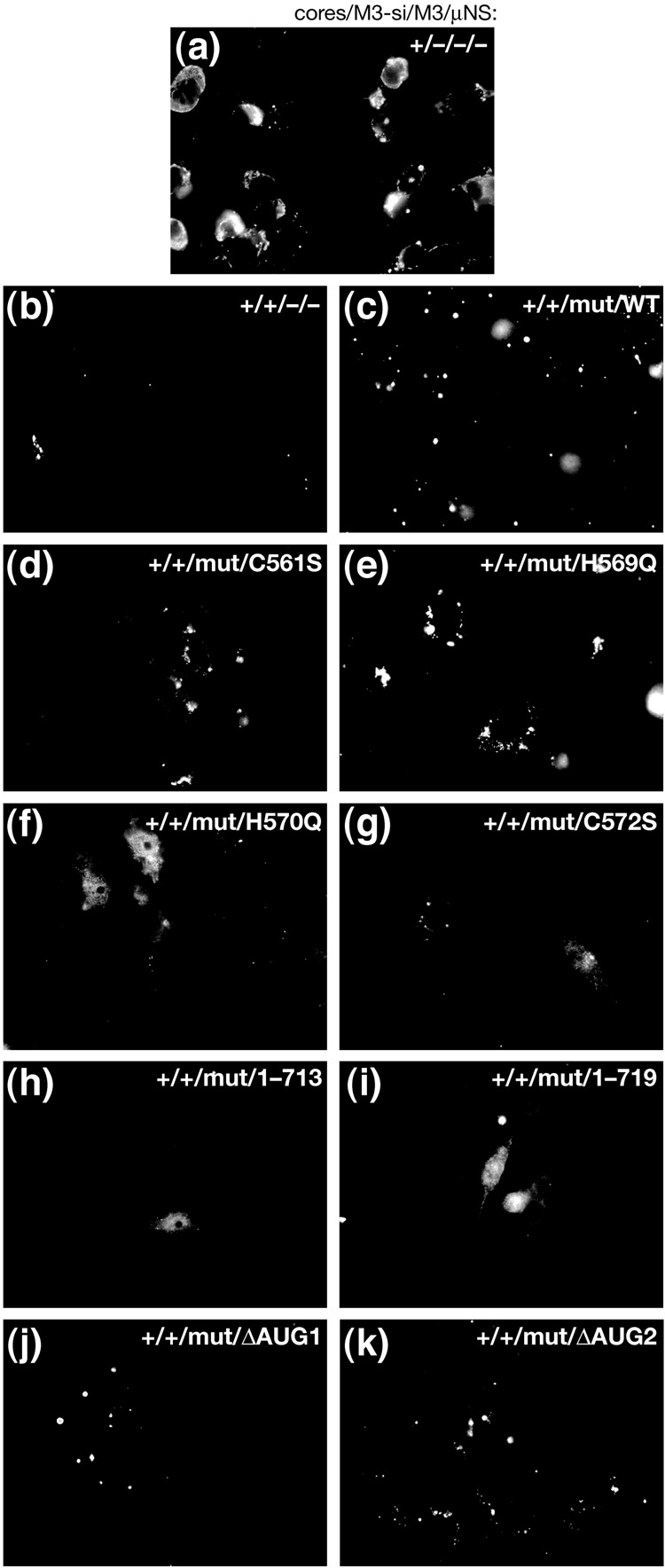
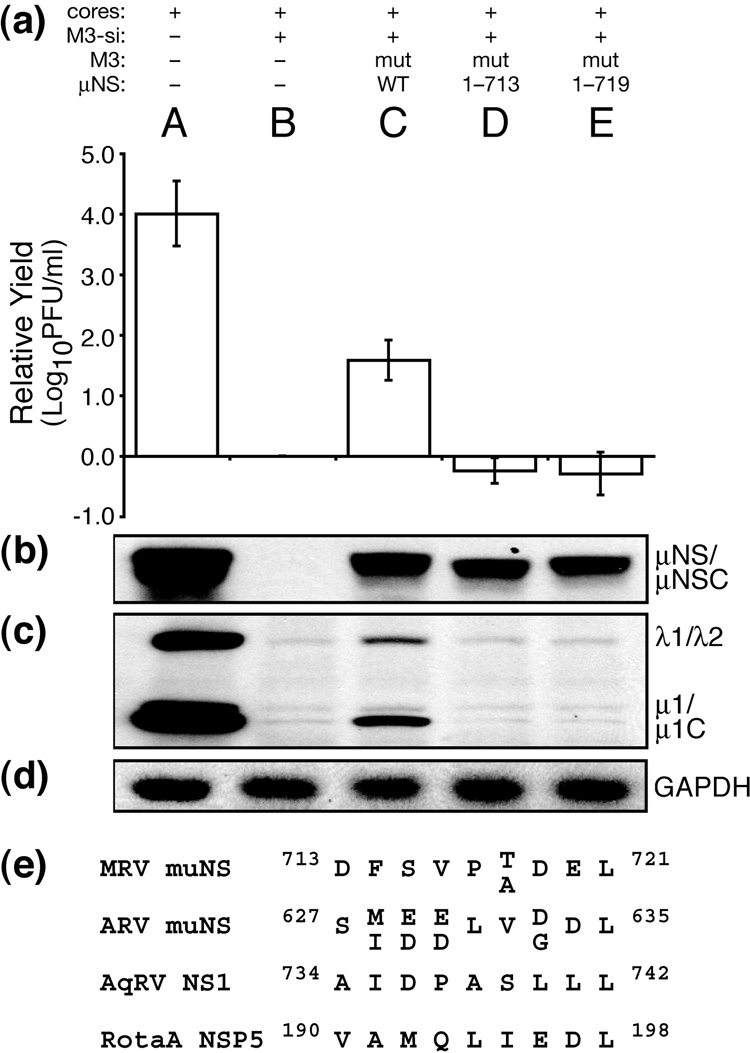
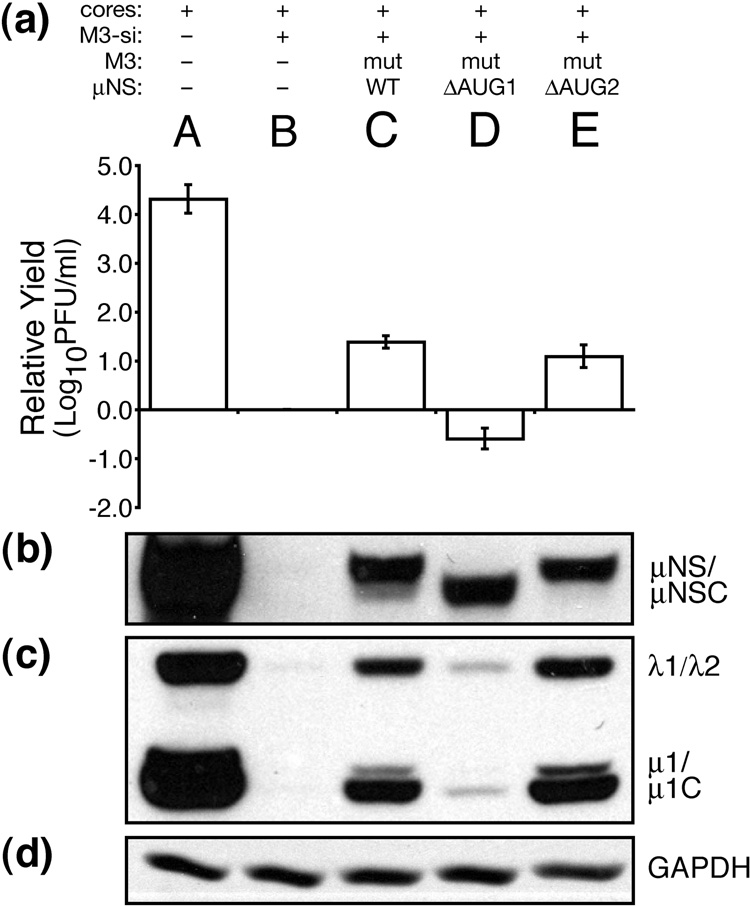
Genome replication of mammalian orthoreovirus (MRV) occurs in cytoplasmic inclusion bodies called viral factories. Nonstructural protein microNS, encoded by genome segment M3, is a major constituent of these structures. When expressed without other viral proteins, microNS forms cytoplasmic inclusions morphologically similar to factories, suggesting a role for microNS as the factory framework or matrix. In addition, most other MRV proteins, including all five core proteins (lambda1, lambda2, lambda3, micro2, and sigma2) and nonstructural protein sigmaNS, can associate with microNS in these structures. In the current study, small interfering RNA targeting M3 was transfected in association with MRV infection and shown to cause a substantial reduction in microNS expression as well as, among other effects, a reduction in infectious yields by as much as 4 log(10) values. By also transfecting in vitro-transcribed M3 plus-strand RNA containing silent mutations that render it resistant to the small interfering RNA, we were able to complement microNS expression and to rescue infectious yields by ~100-fold. We next used microNS mutants specifically defective at forming factory-matrix structures to show that this function of microNS is important for MRV growth; point mutations in a C-proximal, putative zinc-hook motif as well as small deletions at the extreme C terminus of microNS prevented rescue of viral growth while causing microNS to be diffusely distributed in cells. We furthermore confirmed that an N-terminally truncated form of microNS, designed to represent microNSC and still able to form factory-matrix structures, is unable to rescue MRV growth, localizing one or more other important functions to an N-terminal region of microNS known to be involved in both micro2 and sigmaNS association. Thus, factory-matrix formation is an important, though not a sufficient function of microNS during MRV infection; microNS is multifunctional in the course of viral growth.
Figures
References
-
- Ahmed R, Canning WM, Kauffman RS, Sharpe AH, Hallum JV, Fields BN. Role of the host cell in persistent viral infection: coevolution of L cells and reovirus during persistent infection. Cell. 1981;25:325–332. - PubMed
-
- Astell C, Silverstein SC, Levin DH, Acs G. Regulation of the reovirus RNA transcriptase by a viral capsomere protein. Virology. 1972;48:648–654. - PubMed
-
- Attoui H, Fang Q, Mohd Jaafar F, Cantaloube JF, Biagini P, de Micco P, de Lamballerie X. Common evolutionary origin of aquareoviruses and orthoreoviruses revealed by genome characterization of Golden shiner reovirus, Grass carp reovirus, Striped bass reovirus and golden ide reovirus (genus Aquareovirus, family Reoviridae) J. Gen. Virol. 2002;83:1941–1951. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
