

Transcriptional analysis and functional characterization of a gene pair encoding iron-regulated xenocin and immunity proteins of Xenorhabdus nematophila
- PMID: 18375563
- PMCID: PMC2395030
- DOI: 10.1128/JB.00209-08
Transcriptional analysis and functional characterization of a gene pair encoding iron-regulated xenocin and immunity proteins of Xenorhabdus nematophila
Abstract
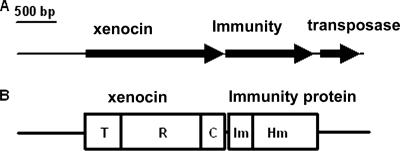
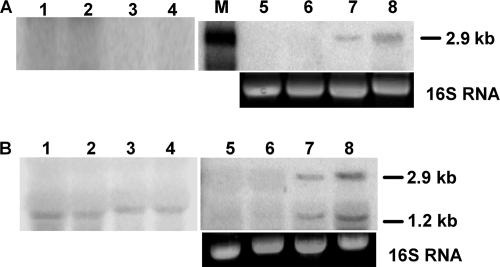
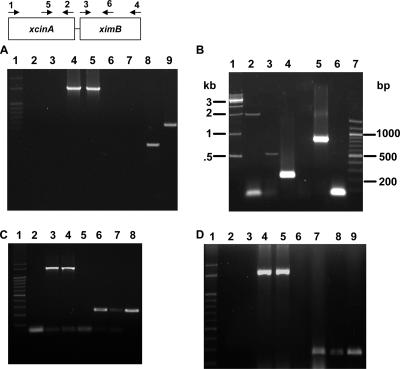
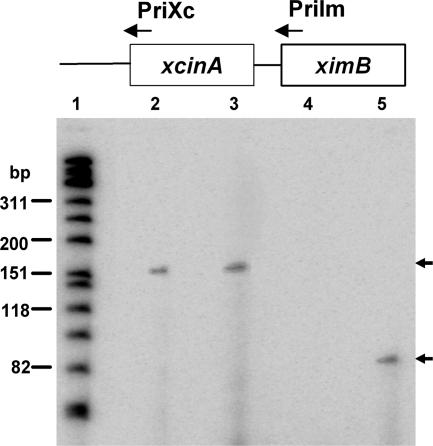
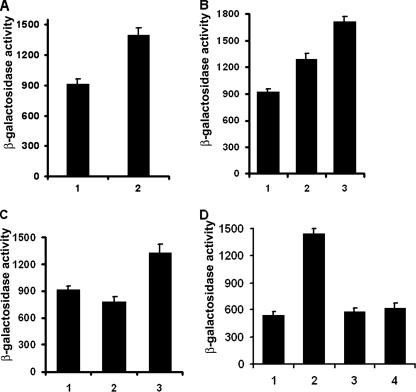
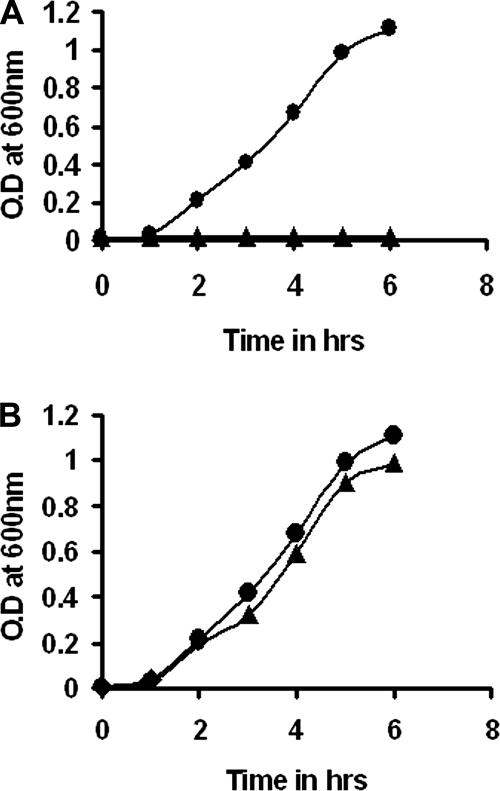
We describe a two-gene cluster encoding a bacteriocin, xenocin, and the cognate immunity protein in the insect-pathogenic bacterium Xenorhabdus nematophila, which infects and kills larval stages of the common crop pest Helicoverpa armigera. The two genes, xcinA and ximB, are present in the genome as a single transcriptional unit, which is regulated under SOS conditions. The stress-inducible promoter was activated by mitomycin C, glucose, and Fe(3+) depletion and at an elevated temperature when it was tested in Escherichia coli cells. Expression of the xenocin protein alone in E. coli inhibited the growth of this organism. The growth inhibition was abolished when the immunity protein was also present. A recombinant xenocin-immunity protein complex inhibited the growth of E. coli indicator cells when it was added exogenously to a growing culture. Xenocin is an endoribonuclease with an enzymatically active C-terminal domain. Six resident bacterial species (i.e., Bacillus, Enterobacter, Enterococcus, Citrobacter, Serratia, and Stenotrophomonas species) from the H. armigera gut exhibited sensitivity to recombinant xenocin when the organisms were grown under iron-depleted conditions and at a high temperature. Xenocin also inhibited the growth of two Xenorhabdus isolates. This study demonstrates that Fe(3+) depletion acts as a common cue for synthesis of xenocin by X. nematophila and sensitization of the target strains to the bacteriocin.
Figures

References
-
- Akhurst, R. J. 1982. Antibiotic activity of Xenorhabdus spp. bacteria symbiotically associated with insect pathogenic nematodes of the families Heterorhabditae and Steinernematidae. J. Gen. Microbiol. 1283061-3065. - PubMed
-
- Akhurst, R. J., and G. B. Dunphy. 1993. Symbiotically associated entomopathogenic bacteria, nematodes and their insect hosts, p. 1-23. In N. Beckage, S. Thompson, and B. Federici (ed.), Parasites and pathogens of insects, vol. 2. Academic Press, Inc., New York, NY.
-
- Ausbel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1989. Current protocols in molecular biology. John Wiley and Sons, New York, NY.
-
- Barnard, T. J., M. E. Watson, Jr., and M. A. McIntosh. 2001. Mutations in the Escherichia coli receptor FepA reveal residues involved in ligand binding and transport. Mol. Microbiol. 41527-536. - PubMed
-
- Bedding, R. A., and R. J. Akhrust. 1975. Nematodes and their biological control of insect pests. Nematologia 21109-110.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
