

Tumor immunotherapy across MHC barriers using allogeneic T-cell precursors
- PMID: 18376399
- PMCID: PMC2731996
- DOI: 10.1038/nbt1395
Tumor immunotherapy across MHC barriers using allogeneic T-cell precursors
Erratum in
-
Author Correction: Tumor immunotherapy across MHC barriers using allogeneic T-cell precursors.Nat Biotechnol. 2024 Apr;42(4):674. doi: 10.1038/s41587-024-02205-3. Nat Biotechnol. 2024. PMID: 38531977 No abstract available.
Abstract
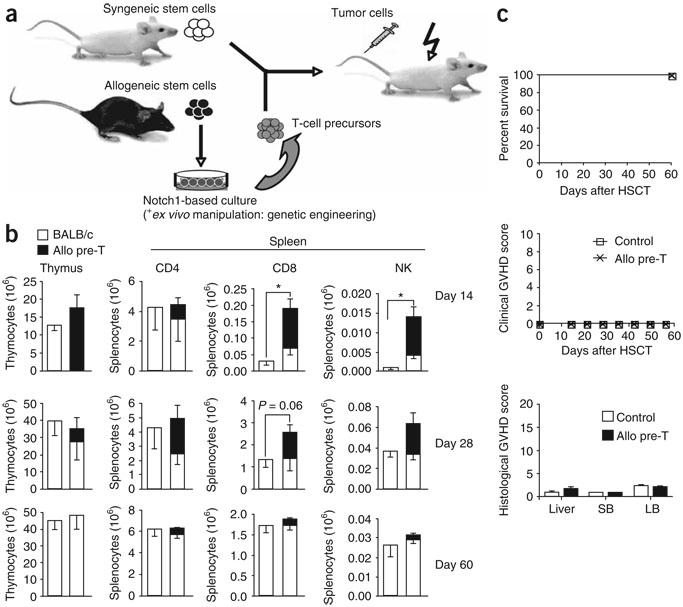
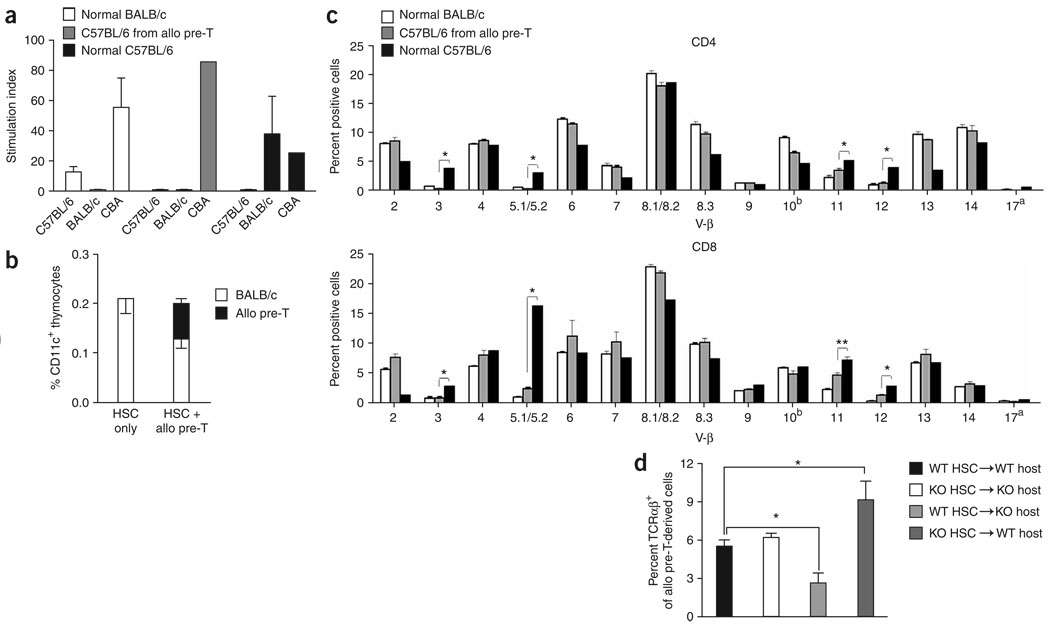
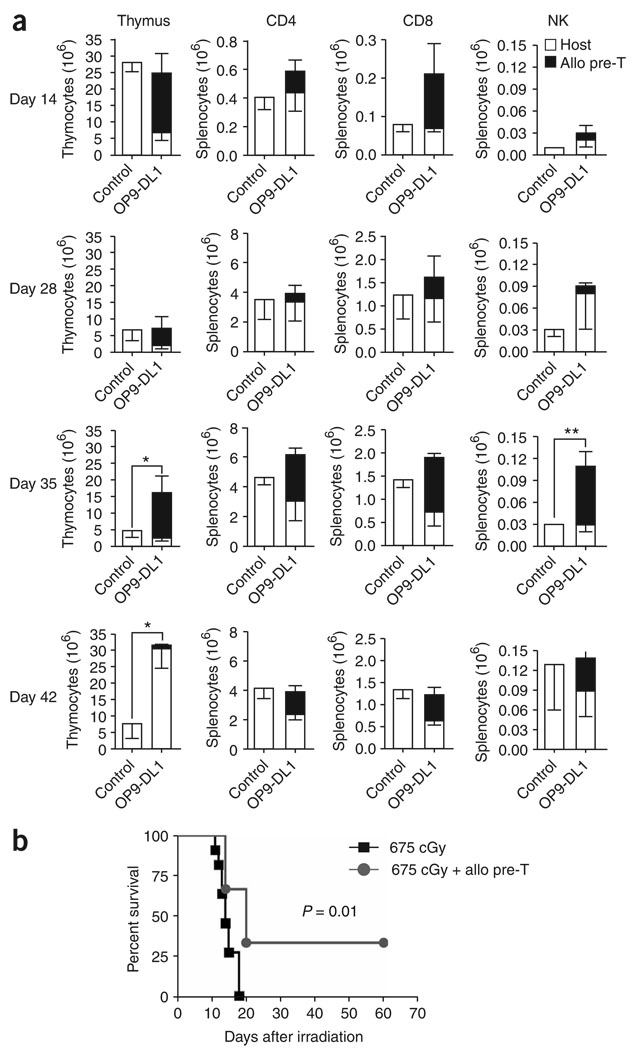
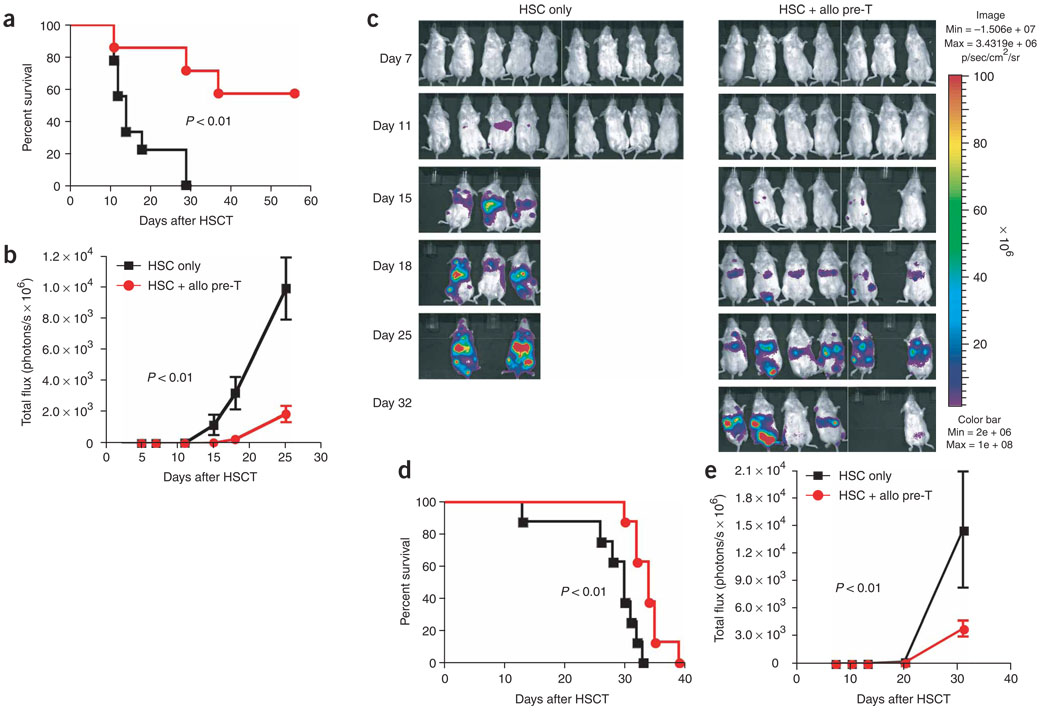
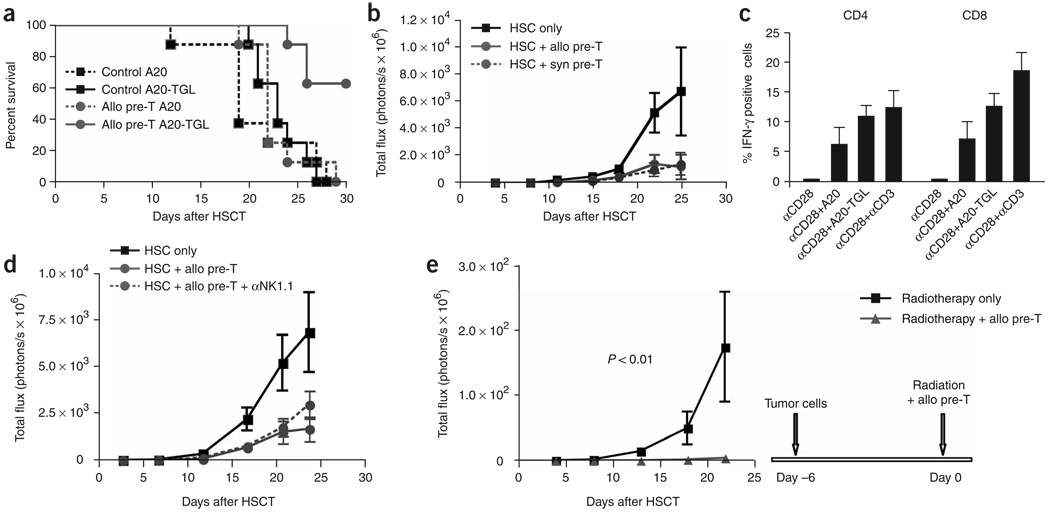
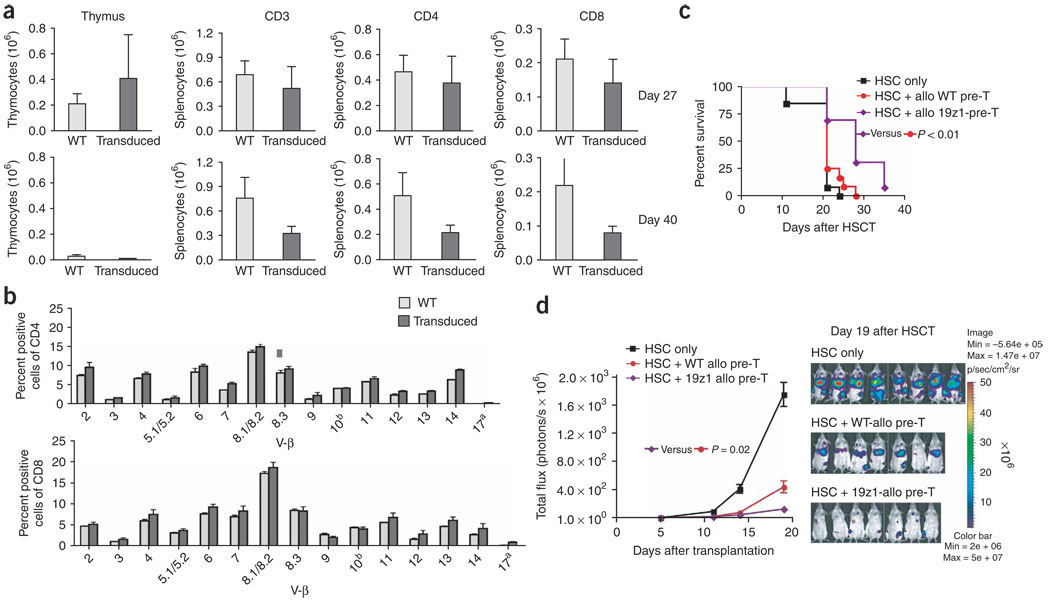
We present a strategy for adoptive immunotherapy using T-lineage committed lymphoid precursor cells generated by Notch1-based culture. We found that allogeneic T-cell precursors can be transferred to irradiated individuals irrespective of major histocompatibility complex (MHC) disparities and give rise to host-MHC restricted and host-tolerant functional allogeneic T cells, improving survival in irradiated recipients as well as enhancing anti-tumor responses. T-cell precursors transduced to express a chimeric receptor targeting hCD19 resulted in significant additional anti-tumor activity, demonstrating the feasibility of genetic engineering of these cells. We conclude that ex vivo generated MHC-disparate T-cell precursors from any donor can be used universally for 'off-the-shelf' immunotherapy, and can be further enhanced by genetic engineering for targeted immunotherapy.
Figures
References
-
- Mackall CL, Gress RE. Thymic aging and T-cell regeneration. Immunol. Rev. 1997;160:91–102. - PubMed
-
- Grunebaum E, Sharfe N, Roifman CM. Human T cell immunodeficiency: when signal transduction goes wrong. Immunol. Res. 2006;35:117–126. - PubMed
-
- Fischer A, et al. Naturally occurring primary deficiencies of the immune system. Annu. Rev. Immunol. 1997;15:93–124. - PubMed
-
- Chinen J, Finkelman F, Shearer WT. Advances in basic and clinical immunology. J. Allergy Clin. Immunol. 2006;118:489–495. - PubMed
-
- Lehrnbecher T, Foster C, Vázquez N, Mackall CL, Chanock SJ. Therapy-induced alterations in host defense in children receiving therapy for cancer. J. Pediatr. Hematol. Oncol. 1997;19:399–417. - PubMed
Publication types
MeSH terms
Grants and funding
- CA107096/CA/NCI NIH HHS/United States
- HL69929/HL/NHLBI NIH HHS/United States
- R01 HL069929/HL/NHLBI NIH HHS/United States
- R01 CA107096/CA/NCI NIH HHS/United States
- GM07739/GM/NIGMS NIH HHS/United States
- P30 CA08748/CA/NCI NIH HHS/United States
- P30 CA008748/CA/NCI NIH HHS/United States
- P01 CA059350/CA/NCI NIH HHS/United States
- CA59350/CA/NCI NIH HHS/United States
- R24 CA83084/CA/NCI NIH HHS/United States
- T32 GM007739/GM/NIGMS NIH HHS/United States
- CA33049/CA/NCI NIH HHS/United States
- R24 CA083084/CA/NCI NIH HHS/United States
- CA40350/CA/NCI NIH HHS/United States
- P01 CA033049/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
