

Direct proteasome-independent cross-presentation of viral antigen by plasmacytoid dendritic cells on major histocompatibility complex class I
- PMID: 18376401
- PMCID: PMC2695657
- DOI: 10.1038/ni.1602
Direct proteasome-independent cross-presentation of viral antigen by plasmacytoid dendritic cells on major histocompatibility complex class I
Abstract
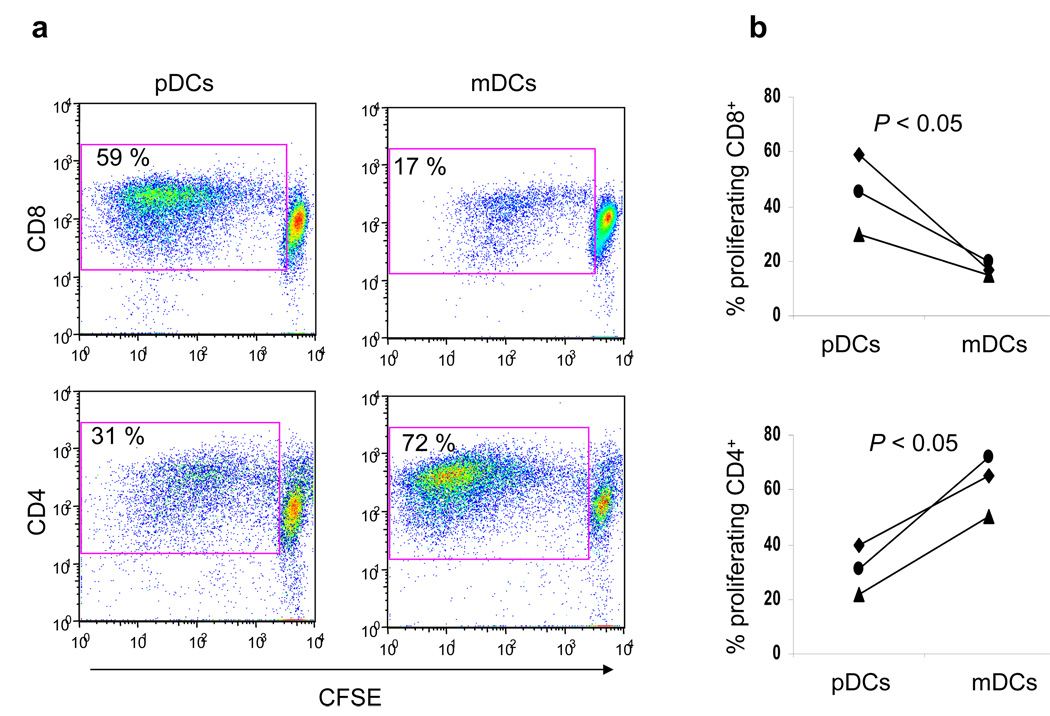
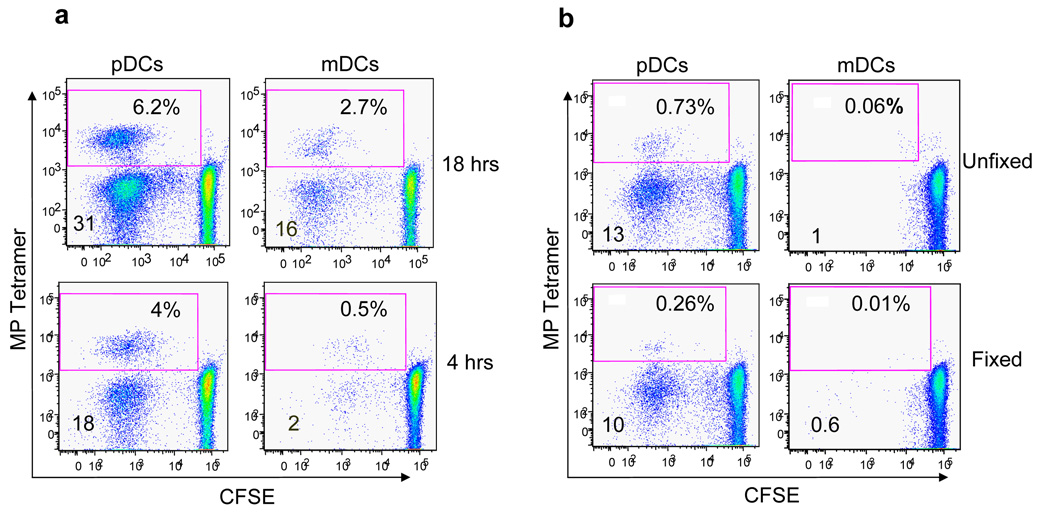
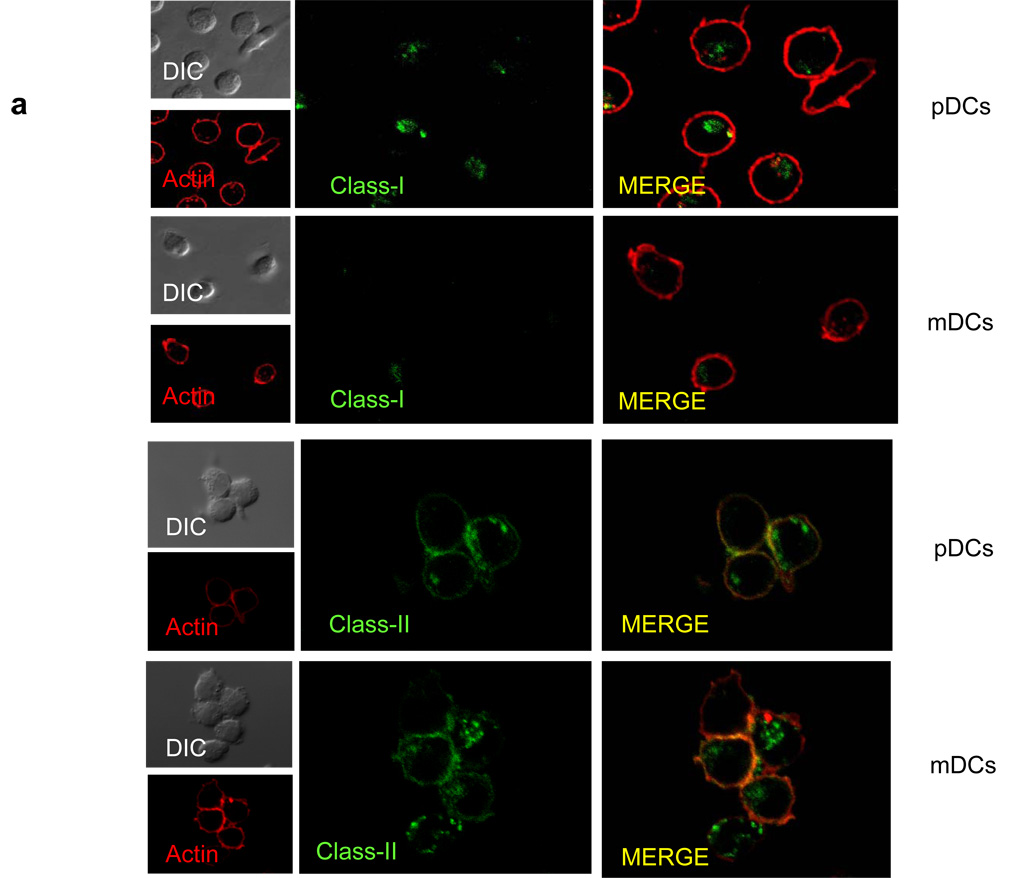
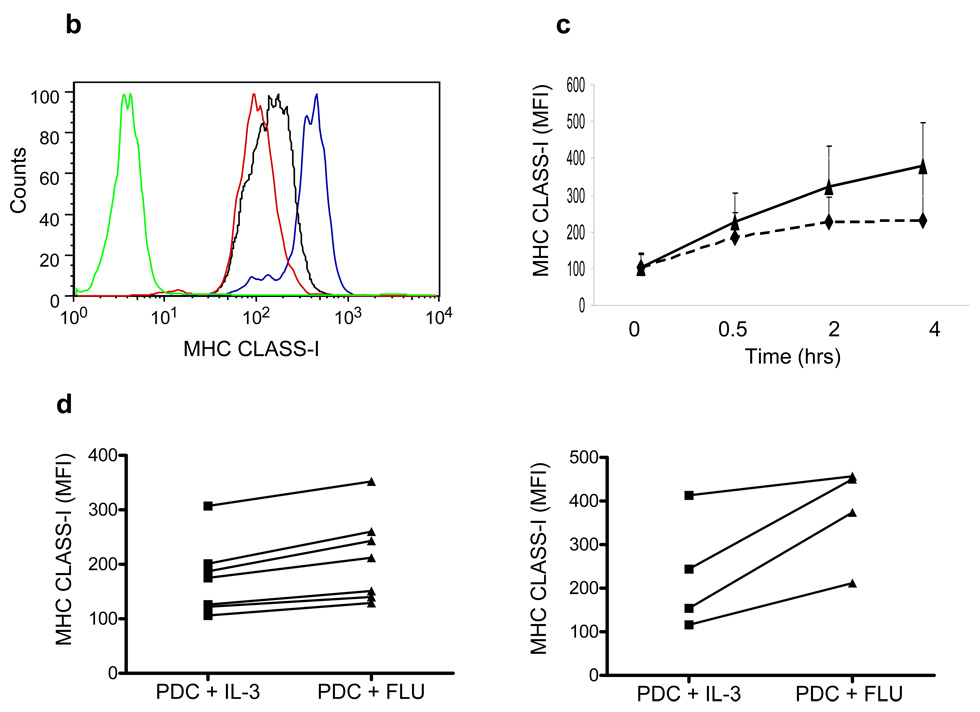
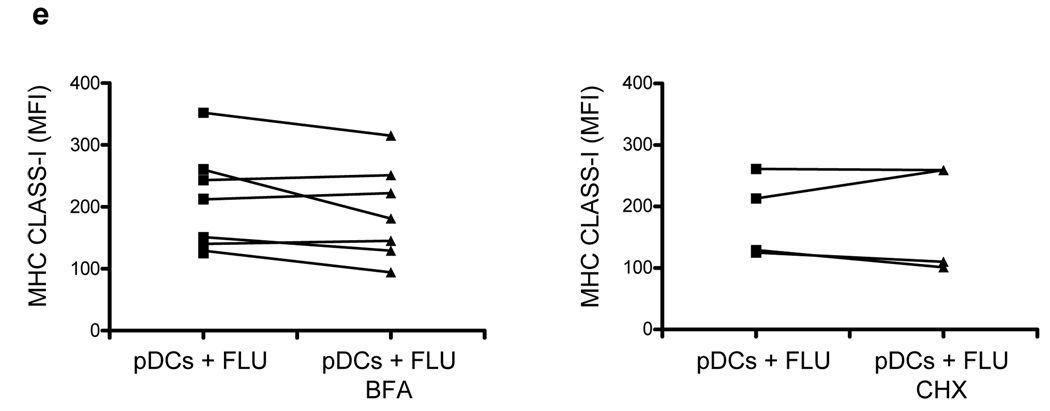
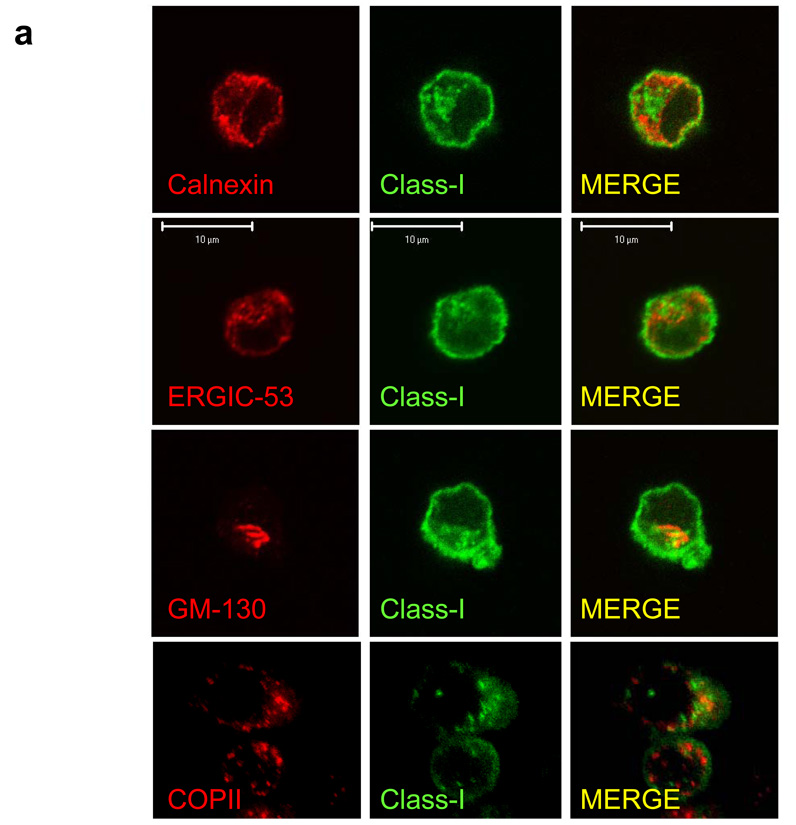
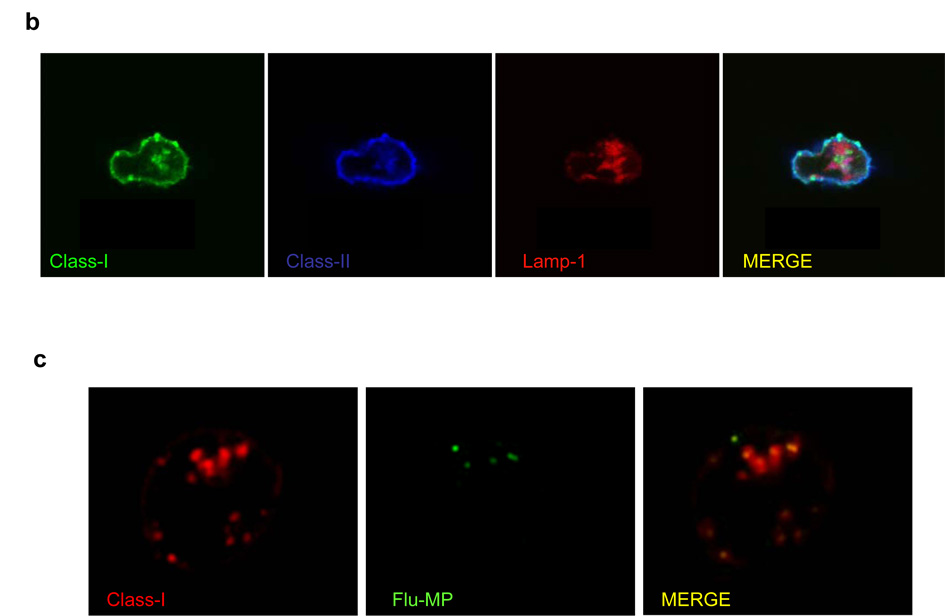
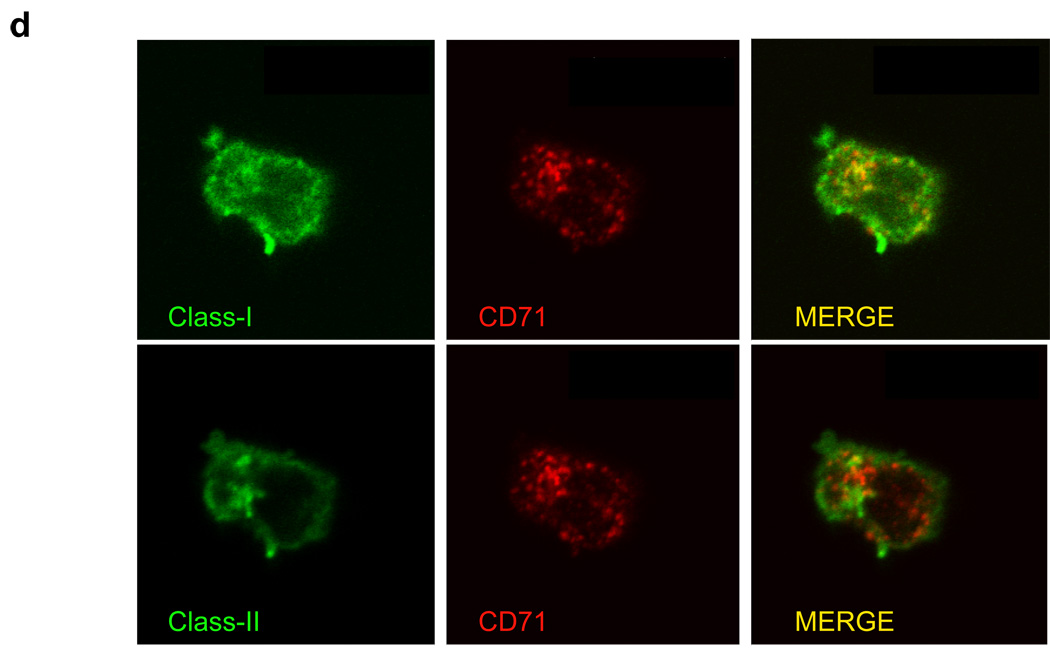
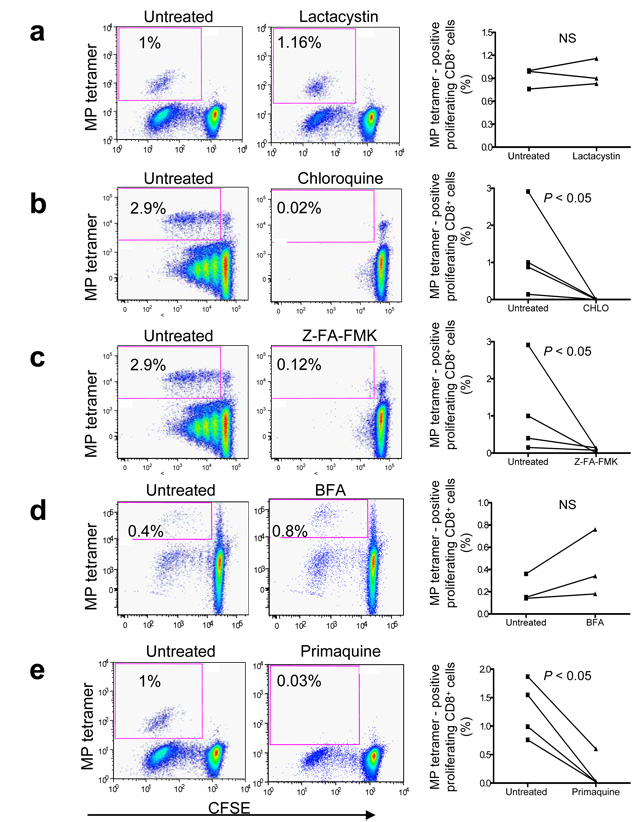
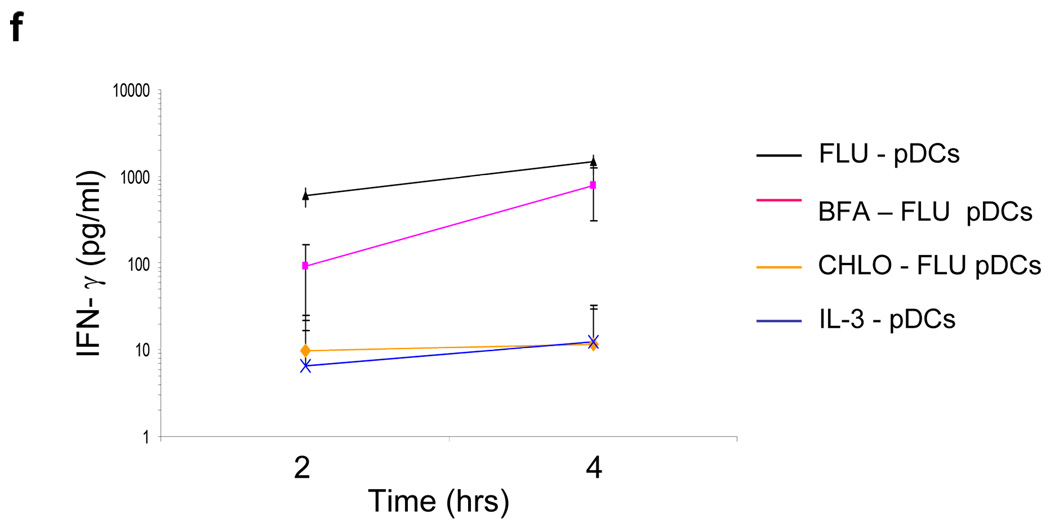
Although plasmacytoid dendritic cells (pDCs) respond to virus replication in a nonspecific way by producing large amounts of type I interferon, a rapid, direct function for pDCs in activating antiviral lymphocytes is less apparent. Here we show that pDCs were able to rapidly initiate antigen-specific antiviral CD8+ T cell responses. After being exposed to virus, pDCs efficiently and rapidly internalized exogenous viral antigens and then presented those antigens on major histocompatibility complex (MHC) class I to CD8+ T cells. Processing of exogenous antigen occurred in endocytic organelles and did not require transit of antigen to the cytosol. Intracellular stores of MHC class I partially localized together with the transferrin receptor and internalized transferrin in endosomes, which suggested that such recycling endosomes are sites for loading peptide onto MHC class I or for peptide transit. Our data demonstrate that pDCs use 'ready-made' stores of MHC class I to rapidly present exogenous antigen to CD8+ T cells.
Figures
Comment in
-
Cross-presentation: avoiding trafficking chaos?Nat Immunol. 2008 May;9(5):461-3. doi: 10.1038/ni0508-461. Nat Immunol. 2008. PMID: 18425099 No abstract available.
References
-
- Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245–252. - PubMed
-
- Shortman K, Liu YJ. Mouse and human dendritic cell subtypes. Nat. Rev. Immunol. 2002;2:151–161. - PubMed
-
- Mellman I, Steinman RM. Dendritic cells: specialized and regulated antigen processing machines. Cell. 2001;106:255–258. - PubMed
-
- Reis ESousa. Dendritic cells in a mature age. Nat. Rev. Immunol. 2006;6:476–483. - PubMed
-
- Watts C. The exogenous pathway for antigen presentation on major histocompatibility complex class II and CD1 molecules. Nat. Immunol. 2004;5:685–692. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
