

Plk1- and beta-TrCP-dependent degradation of Bora controls mitotic progression
- PMID: 18378770
- PMCID: PMC2287288
- DOI: 10.1083/jcb.200712027
Plk1- and beta-TrCP-dependent degradation of Bora controls mitotic progression
Abstract
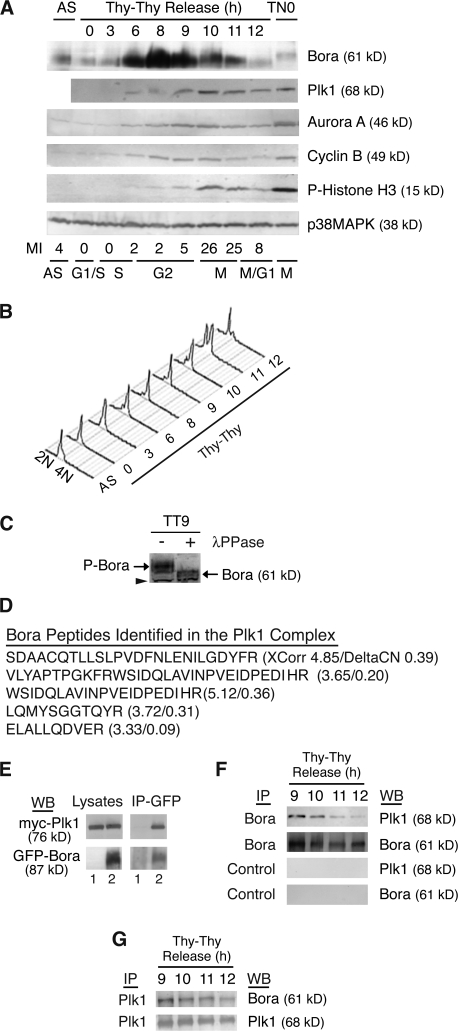
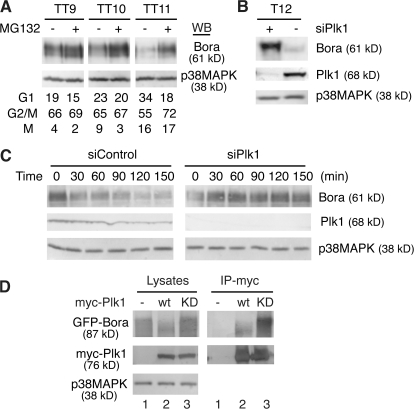
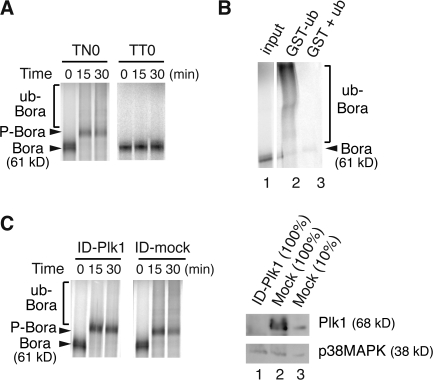
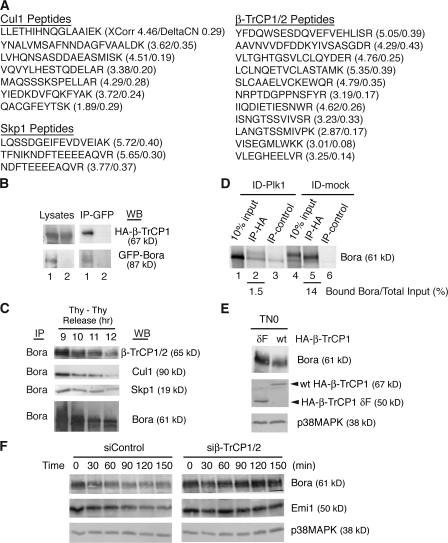
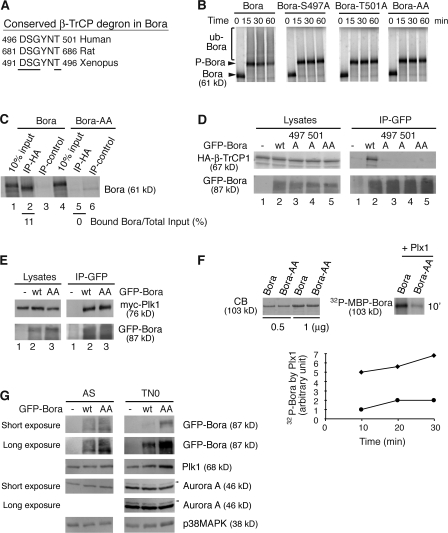
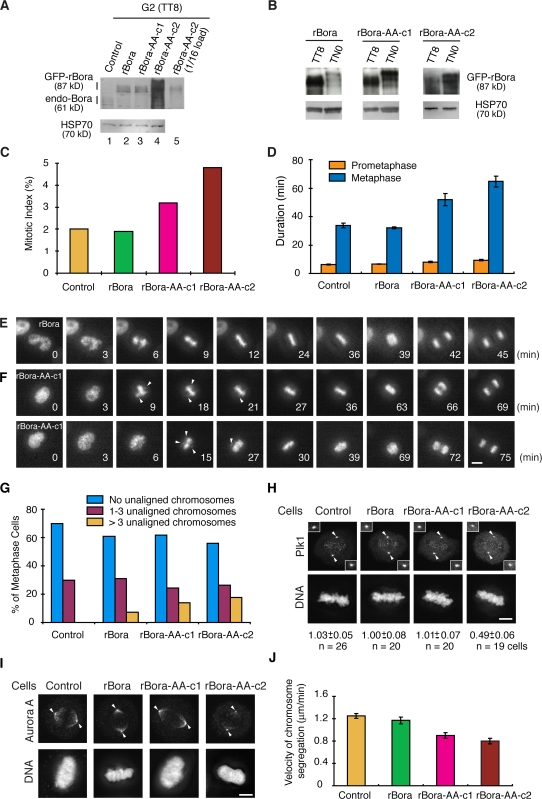
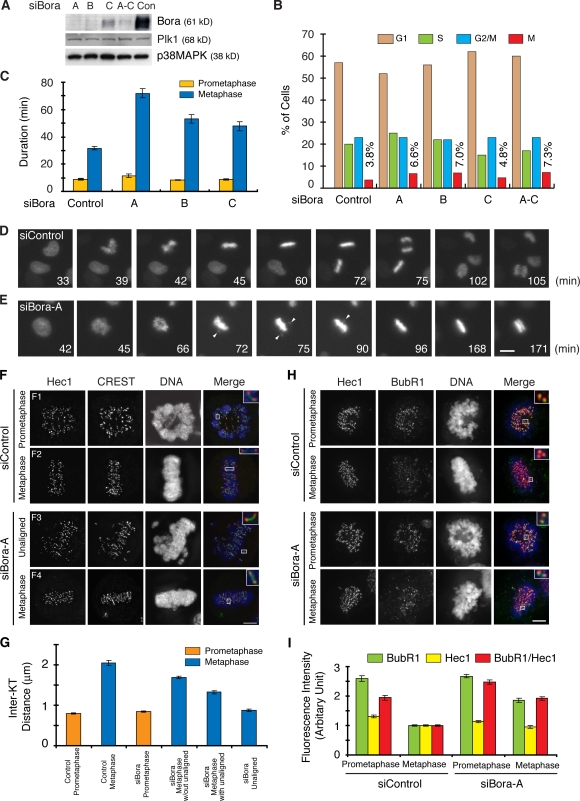
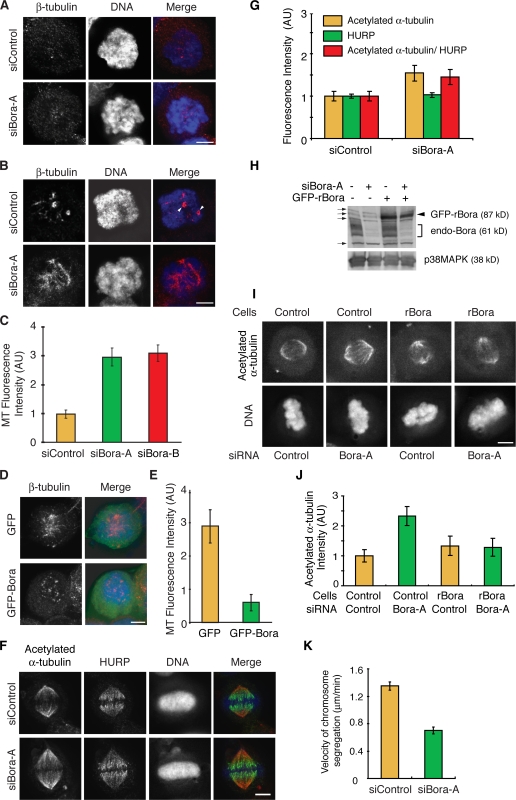
Through a convergence of functional genomic and proteomic studies, we identify Bora as a previously unknown cell cycle protein that interacts with the Plk1 kinase and the SCF-beta-TrCP ubiquitin ligase. We show that the Bora protein peaks in G2 and is degraded by proteasomes in mitosis. Proteolysis of Bora requires the Plk1 kinase activity and is mediated by SCF-beta-TrCP. Plk1 phosphorylates a conserved DSGxxT degron in Bora and promotes its interaction with beta-TrCP. Mutations in this degron stabilize Bora. Expression of a nondegradable Bora variant prolongs the metaphase and delays anaphase onset, indicating a physiological requirement of Bora degradation. Interestingly, the activity of Bora is also required for normal mitotic progression, as knockdown of Bora activates the spindle checkpoint and delays sister chromatid segregation. Mechanistically, Bora regulates spindle stability and microtubule polymerization and promotes tension across sister kinetochores during mitosis. We conclude that tight regulation of the Bora protein by its synthesis and degradation is critical for cell cycle progression.
Figures
Similar articles
-
PLK1 and β-TrCP-dependent ubiquitination and degradation of Rap1GAP controls cell proliferation.PLoS One. 2014 Oct 17;9(10):e110296. doi: 10.1371/journal.pone.0110296. eCollection 2014. PLoS One. 2014. PMID: 25329897 Free PMC article.
-
Polo-like kinase-1 controls proteasome-dependent degradation of Claspin during checkpoint recovery.Curr Biol. 2006 Oct 10;16(19):1950-5. doi: 10.1016/j.cub.2006.08.026. Epub 2006 Aug 24. Curr Biol. 2006. PMID: 16934469
-
Plk1 regulates mitotic Aurora A function through betaTrCP-dependent degradation of hBora.Chromosoma. 2008 Oct;117(5):457-69. doi: 10.1007/s00412-008-0165-5. Epub 2008 Jun 3. Chromosoma. 2008. PMID: 18521620 Free PMC article.
-
The equilibrium of ubiquitination and deubiquitination at PLK1 regulates sister chromatid separation.Cell Mol Life Sci. 2017 Jun;74(12):2127-2134. doi: 10.1007/s00018-017-2457-5. Epub 2017 Feb 10. Cell Mol Life Sci. 2017. PMID: 28188342 Free PMC article. Review.
-
Polo-like kinase 1: target and regulator of anaphase-promoting complex/cyclosome-dependent proteolysis.Cancer Res. 2006 Jul 15;66(14):6895-8. doi: 10.1158/0008-5472.CAN-06-0358. Cancer Res. 2006. PMID: 16849530 Review.
Cited by
-
Radiation pharmacogenomics: a genome-wide association approach to identify radiation response biomarkers using human lymphoblastoid cell lines.Genome Res. 2010 Nov;20(11):1482-92. doi: 10.1101/gr.107672.110. Epub 2010 Oct 5. Genome Res. 2010. PMID: 20923822 Free PMC article.
-
Ubiquitination-mediated degradation of cell cycle-related proteins by F-box proteins.Int J Biochem Cell Biol. 2016 Apr;73:99-110. doi: 10.1016/j.biocel.2016.02.005. Epub 2016 Feb 6. Int J Biochem Cell Biol. 2016. PMID: 26860958 Free PMC article. Review.
-
Alternative functions of core cell cycle regulators in neuronal migration, neuronal maturation, and synaptic plasticity.Neuron. 2009 May 14;62(3):312-26. doi: 10.1016/j.neuron.2009.03.029. Neuron. 2009. PMID: 19447088 Free PMC article. Review.
-
Calmodulin protects Aurora B on the midbody to regulate the fidelity of cytokinesis.Cell Cycle. 2013 Feb 15;12(4):663-73. doi: 10.4161/cc.23586. Epub 2013 Jan 31. Cell Cycle. 2013. PMID: 23370391 Free PMC article.
-
Regulation of the centrosome cycle.Mol Cell Oncol. 2015 Jul 29;3(2):e1075643. doi: 10.1080/23723556.2015.1075643. eCollection 2016 Mar. Mol Cell Oncol. 2015. PMID: 27308597 Free PMC article. Review.
References
-
- Ang, X.L., and J. Wade Harper. 2005. SCF-mediated protein degradation and cell cycle control. Oncogene. 24:2860–2870. - PubMed
-
- Barr, A.R., and F. Gergely. 2007. Aurora-A: the maker and breaker of spindle poles. J. Cell Sci. 120:2987–2996. - PubMed
-
- Barr, F.A., H.H. Sillje, and E.A. Nigg. 2004. Polo-like kinases and the orchestration of cell division. Nat. Rev. Mol. Cell Biol. 5:429–440. - PubMed
-
- Bennett, L.N., and P.R. Clarke. 2006. Regulation of Claspin degradation by the ubiquitin-proteosome pathway during the cell cycle and in response to ATR-dependent checkpoint activation. FEBS Lett. 580:4176–4181. - PubMed
-
- Bern, M., D. Goldberg, W.H. McDonald, and J.R. Yates III. 2004. Automatic quality assessment of peptide tandem mass spectra. Bioinformatics. 20:I49–I54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
