

Reproductive conflict and the separation of reproductive generations in humans
- PMID: 18378891
- PMCID: PMC2291103
- DOI: 10.1073/pnas.0711911105
Reproductive conflict and the separation of reproductive generations in humans
Abstract
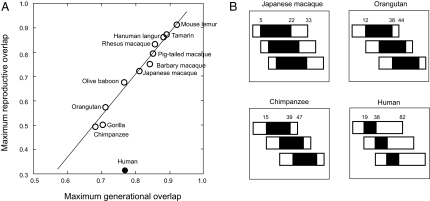
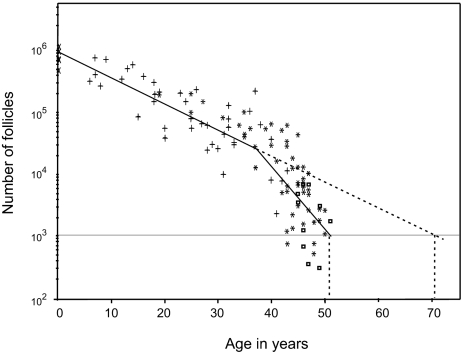
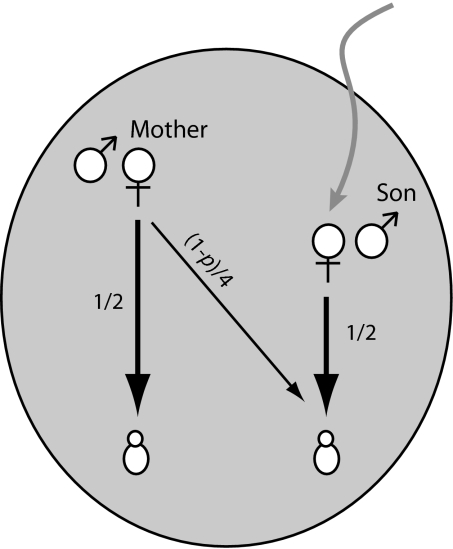
An enduring puzzle of human life history is why women cease reproduction midway through life. Selection can favor postreproductive survival because older females can help their offspring to reproduce. But the kin-selected fitness gains of helping appear insufficient to outweigh the potential benefits of continued reproduction. Why then do women cease reproduction in the first place? Here, we suggest that early reproductive cessation in humans is the outcome of reproductive competition between generations, and we present a simple candidate model of how this competition will be resolved. We show that among primates exhibiting a postreproductive life span, humans exhibit an extraordinarily low degree of reproductive overlap between generations. The rapid senescence of the human female reproductive system coincides with the age at which, in natural fertility populations, women are expected to encounter reproductive competition from breeding females of the next generation. Several lines of evidence suggest that in ancestral hominids, this younger generation typically comprised immigrant females. In these circumstances, relatedness asymmetries within families are predicted to give younger females a decisive advantage in reproductive conflict with older females. A model incorporating both the costs of reproductive competition and the benefits of grandmothering can account for the timing of reproductive cessation in humans and so offers an improved understanding of the evolution of menopause.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Moss CJ. The demography of an African elephant (Loxodonta africana) population in Amboseli, Kenya. J Zool. 2001;255:145–156.
-
- Mizroch SA. Analyses of some biological parameters of the Antarcticfin whale (Balaenopter physalus) Rep Int Whaling Comm. 1981;31:425–434.
-
- Wood JW. Dynamics of Human Reproduction. New York: de Gruyter; 1994.
-
- Blurton-Jones NG, Hawkes K, O'Connell JF. Antiquity of postreproductive life: Are there modern impacts on hunter–gatherer postreproductive life spans? Am J Hum Biol. 2002;14:184–205. - PubMed
-
- Williams GC. Pleiotropy, natural selection, and the evolution of senescence. Evolution (Lawrence, Kans) 1957;11:398–411.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
