

MicroRNA-206: the skeletal muscle-specific myomiR
- PMID: 18381085
- PMCID: PMC2656394
- DOI: 10.1016/j.bbagrm.2008.03.001
MicroRNA-206: the skeletal muscle-specific myomiR
Abstract
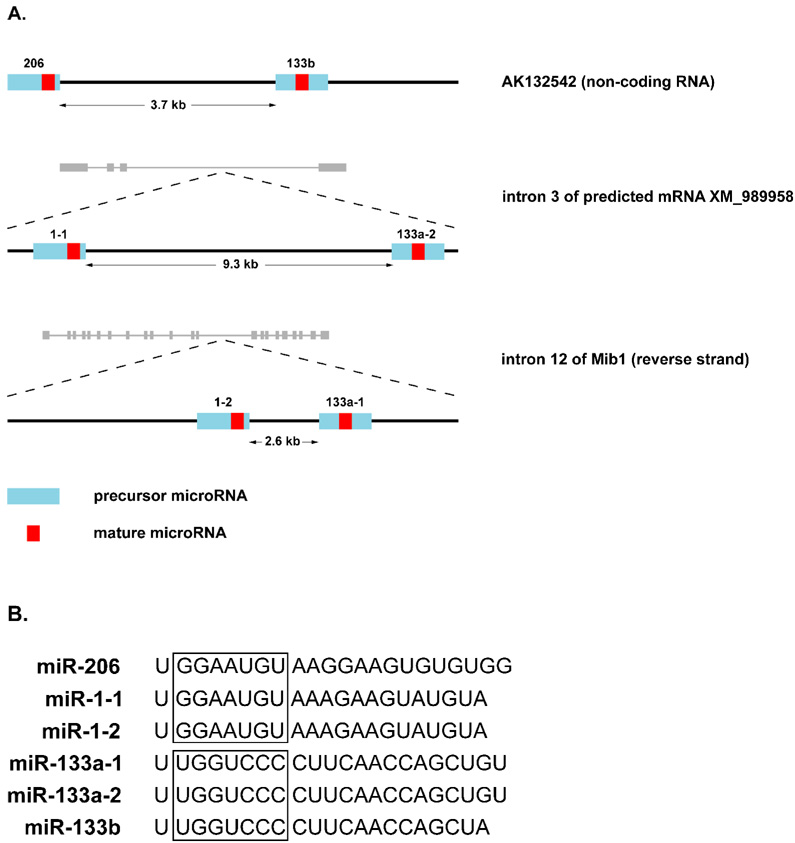
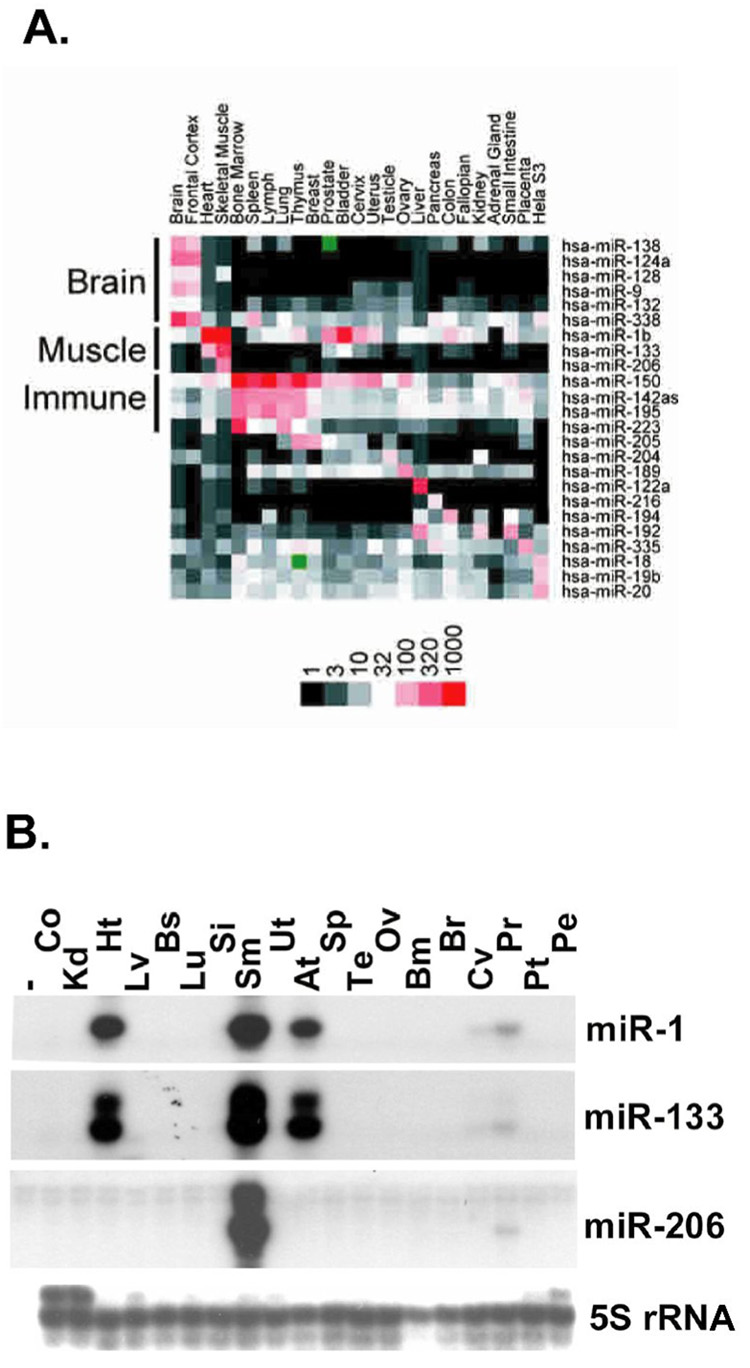
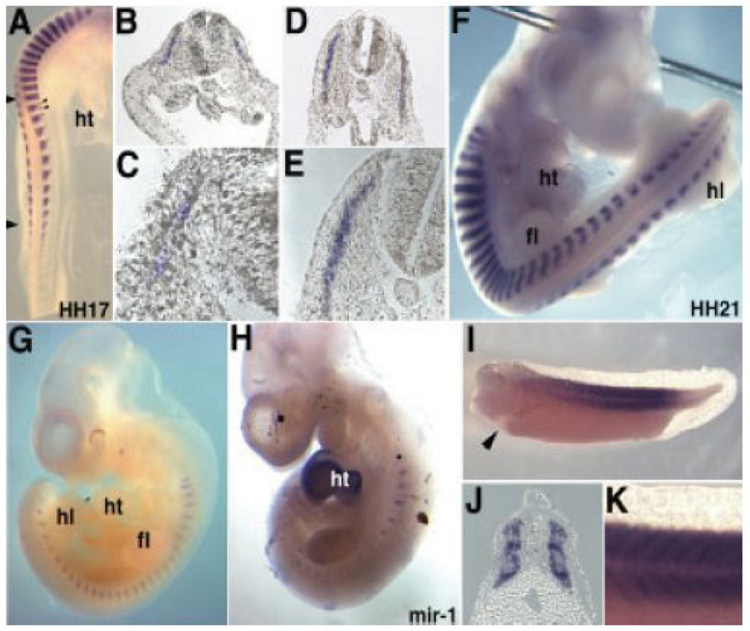
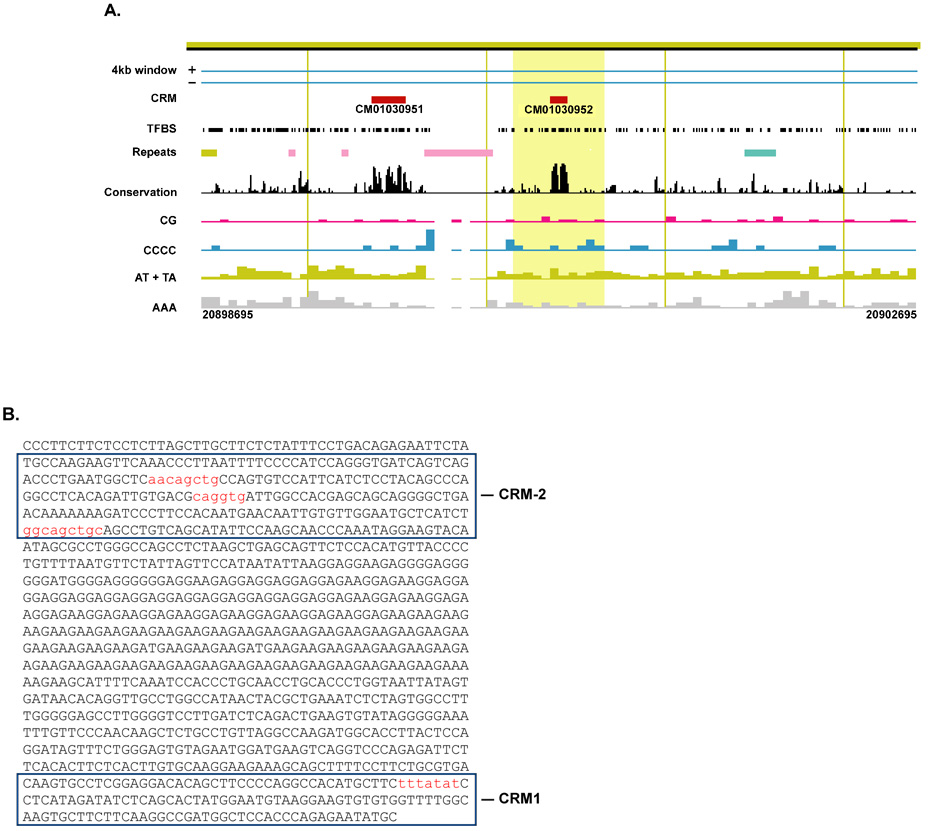
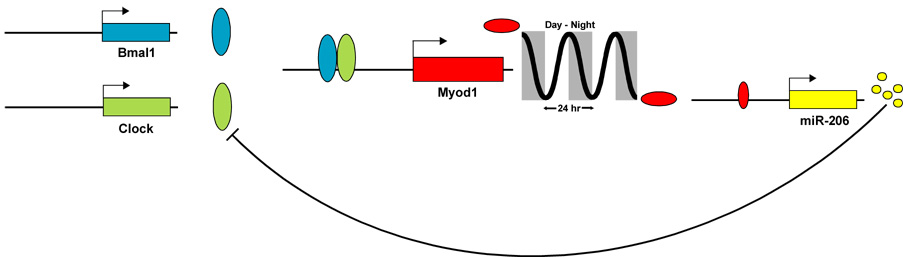
MicroRNAs (miRNAs) are a class of non-coding RNAs involved in post-transcriptional gene silencing. A small number of striated muscle-specific miRNAs have been identified and shown to have an important role in myogenesis, embryonic muscle growth and cardiac function and hypertrophy. One of these myomiRs (myo=muscle+miR=miRNA), miR-206, is unique in that it is only expressed in skeletal muscle. The purpose of this review is to discuss what is currently known about miR-206 and its function in myogenesis as well as propose potential new roles for miR-206 in skeletal muscle biology. The review is also intended to serve as a comprehensive resource for miR-206 with the hope of encouraging further research on the role of miR-206 in skeletal muscle.
Figures
References
-
- Pasquinelli AE, Reinhart BJ, Slack F, Martindale MQ, Kuroda MI, Maller B, Hayward DC, Ball EE, Degnan B, Muller P, Spring J, Srinivasan A, Fishman M, Finnerty J, Corbo J, Levine M, Leahy P, Davidson E, Ruvkun G. Conservation of the sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature. 2000;408:86–89. - PubMed
-
- Arasu P, Wightman B, Ruvkun G. Temporal regulation of lin-14 by the antagonistic action of two other heterochronic genes, lin-4 and lin-28. Genes Dev. 1991;5:1825–1833. - PubMed
-
- Wightman B, Burglin TR, Gatto J, Arasu P, Ruvkun G. Negative regulatory sequences in the lin-14 3'-untranslated region are necessary to generate a temporal switch during Caenorhabditis elegans development. Genes Dev. 1991;5:1813–1824. - PubMed
-
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell. 1993;75:855–862. - PubMed
-
- Ha I, Wightman B, Ruvkun G. A bulged lin-4/lin-14 RNA duplex is sufficient for Caenorhabditis elegans lin-14 temporal gradient formation. Genes Dev. 1996;10:3041–3050. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
