

Cell death by SecTRAPs: thioredoxin reductase as a prooxidant killer of cells
- PMID: 18382651
- PMCID: PMC2268967
- DOI: 10.1371/journal.pone.0001846
Cell death by SecTRAPs: thioredoxin reductase as a prooxidant killer of cells
Abstract
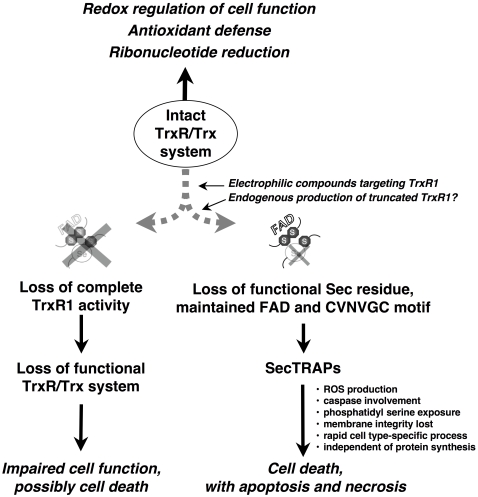
Background: SecTRAPs (selenium compromised thioredoxin reductase-derived apoptotic proteins) can be formed from the selenoprotein thioredoxin reductase (TrxR) by targeting of its selenocysteine (Sec) residue with electrophiles, or by its removal through C-terminal truncation. SecTRAPs are devoid of thioredoxin reductase activity but can induce rapid cell death in cultured cancer cell lines by a gain of function.
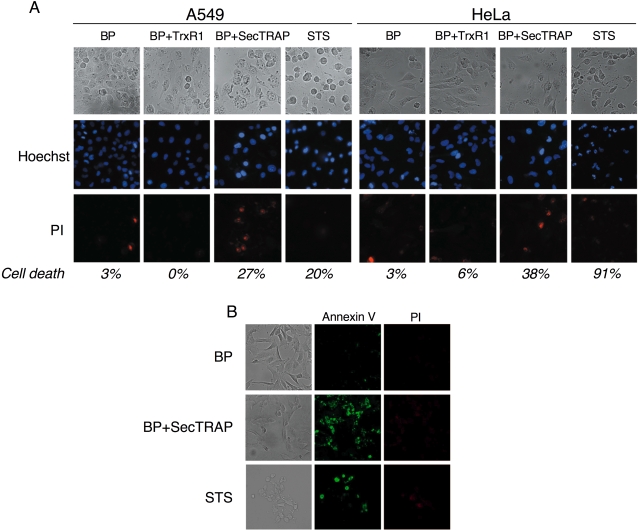
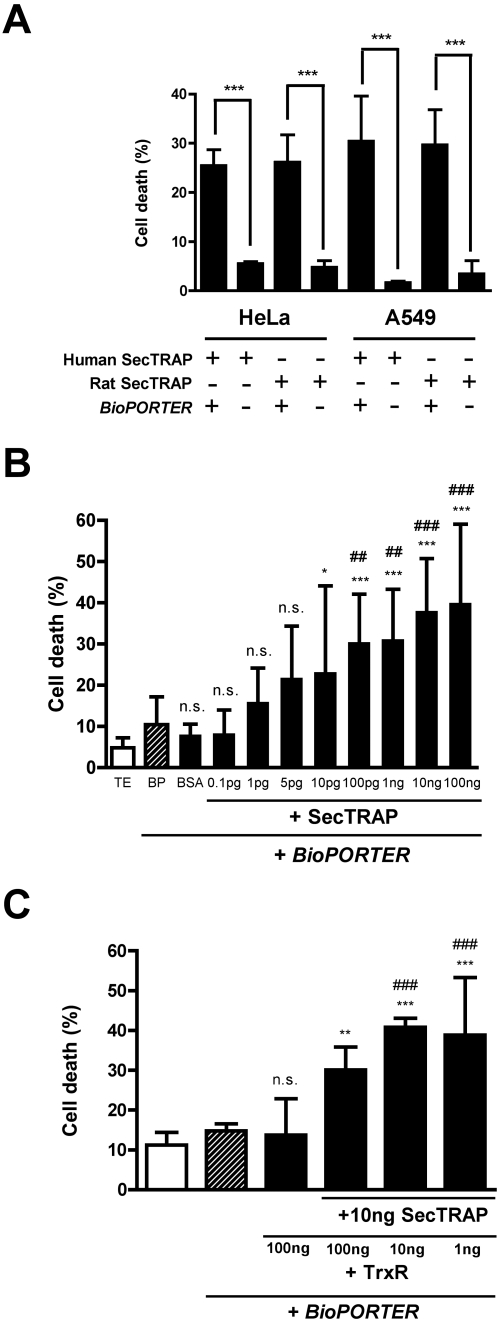
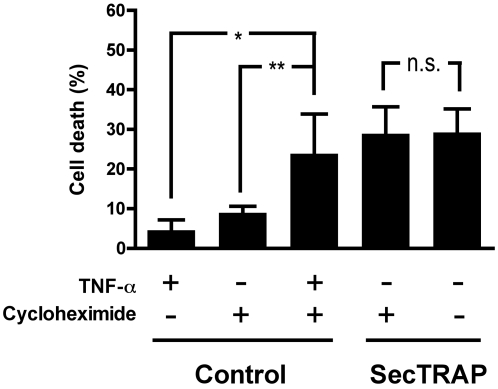
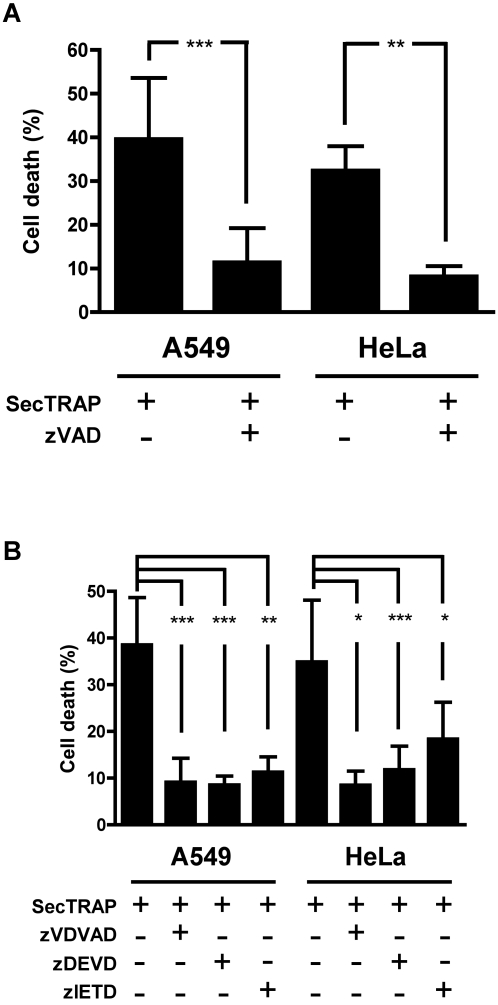
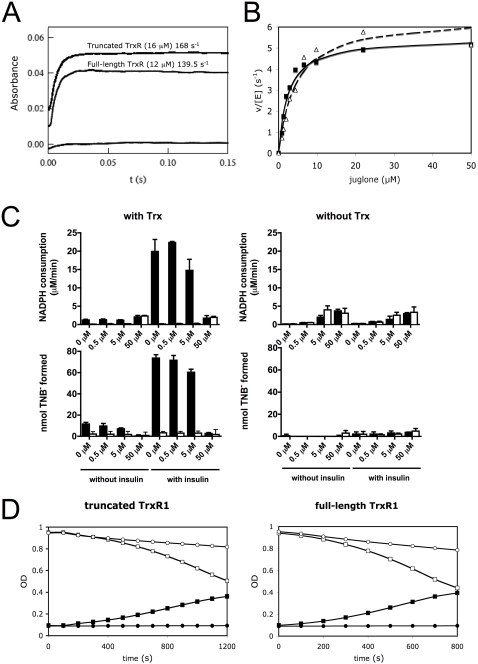
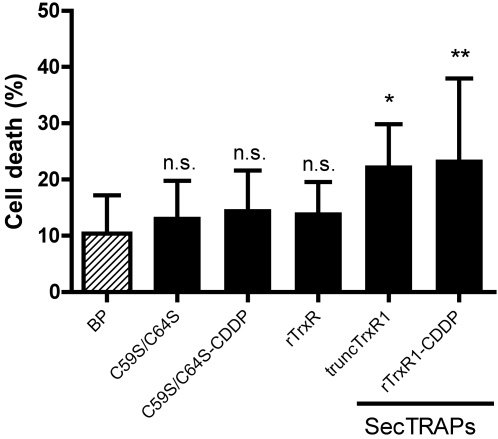
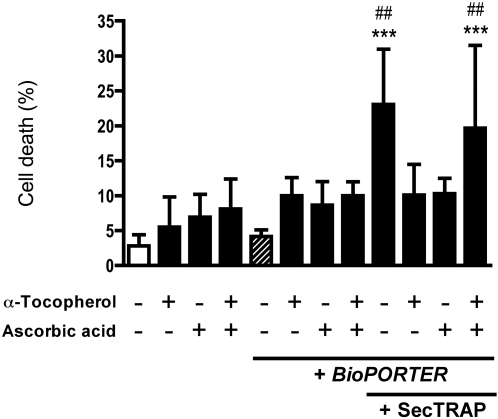
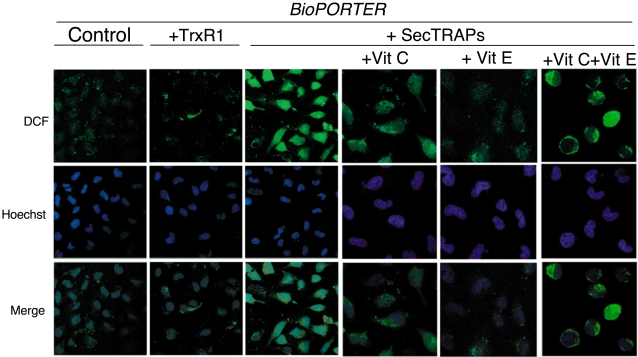
Principal findings: Both human and rat SecTRAPs killed human A549 and HeLa cells. The cell death displayed both apoptotic and necrotic features. It did not require novel protein synthesis nor did it show extensive nuclear fragmentation, but it was attenuated by use of caspase inhibitors. The redox active disulfide/dithiol motif in the N-terminal domain of TrxR had to be maintained for manifestation of SecTRAP cytotoxicity. Stopped-flow kinetics showed that NADPH can reduce the FAD moiety in SecTRAPs at similar rates as in native TrxR and purified SecTRAPs could maintain NADPH oxidase activity, which was accelerated by low molecular weight substrates such as juglone. In a cellular context, SecTRAPs triggered extensive formation of reactive oxygen species (ROS) and consequently antioxidants could protect against the cell killing by SecTRAPs.
Conclusions: We conclude that formation of SecTRAPs could contribute to the cytotoxicity seen upon exposure of cells to electrophilic agents targeting TrxR. SecTRAPs are prooxidant killers of cells, triggering mechanisms beyond those of a mere loss of thioredoxin reductase activity.
Conflict of interest statement
Figures
References
-
- Kryukov GV, Castellano S, Novoselov SV, Lobanov AV, Zehtab O, et al. Characterization of mammalian selenoproteomes. Science. 2003;300:1439–1443. - PubMed
-
- Zhong L, Arnér ESJ, Ljung J, Åslund F, Holmgren A. Rat and calf thioredoxin reductase are homologous to glutathione reductase with a carboxyl-terminal elongation containing a conserved catalytically active penultimate selenocysteine residue. J Biol Chem. 1998;273:8581–8591. - PubMed
-
- Lillig CH, Holmgren A. Thioredoxin and related molecules-from biology to health and disease. Antioxid Redox Signal. 2007;9:25–47. - PubMed
-
- Arnér ESJ, Holmgren A. Physiological functions of thioredoxin and thioredoxin reductase. Eur J Biochem. 2000;267:6102–6109. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
