

Distinct roles of jasmonates and aldehydes in plant-defense responses
- PMID: 18382679
- PMCID: PMC2271129
- DOI: 10.1371/journal.pone.0001904
Distinct roles of jasmonates and aldehydes in plant-defense responses
Abstract
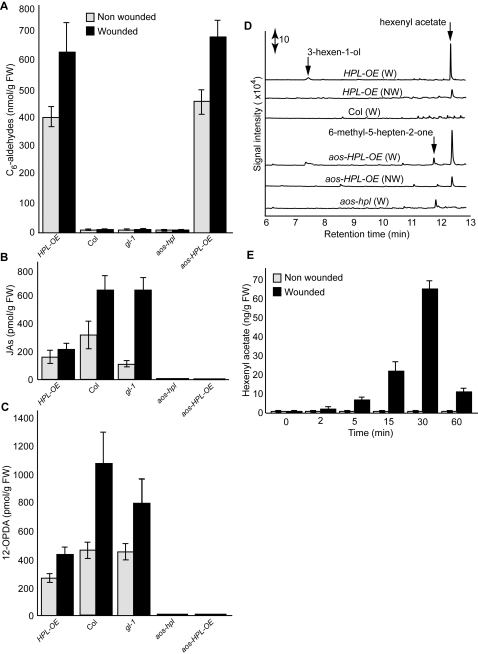
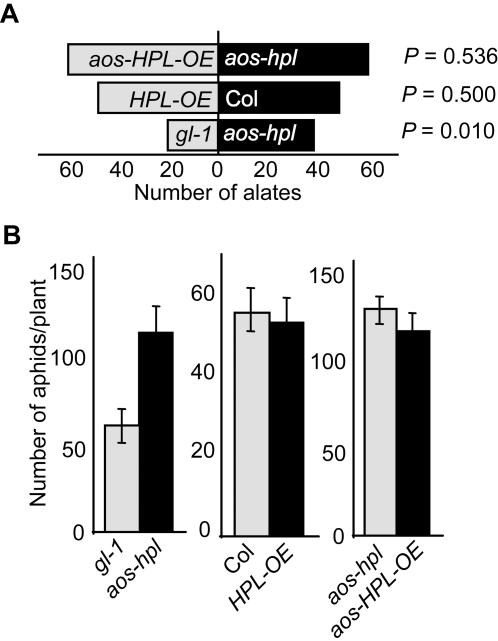
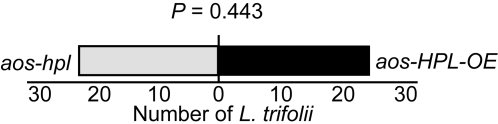
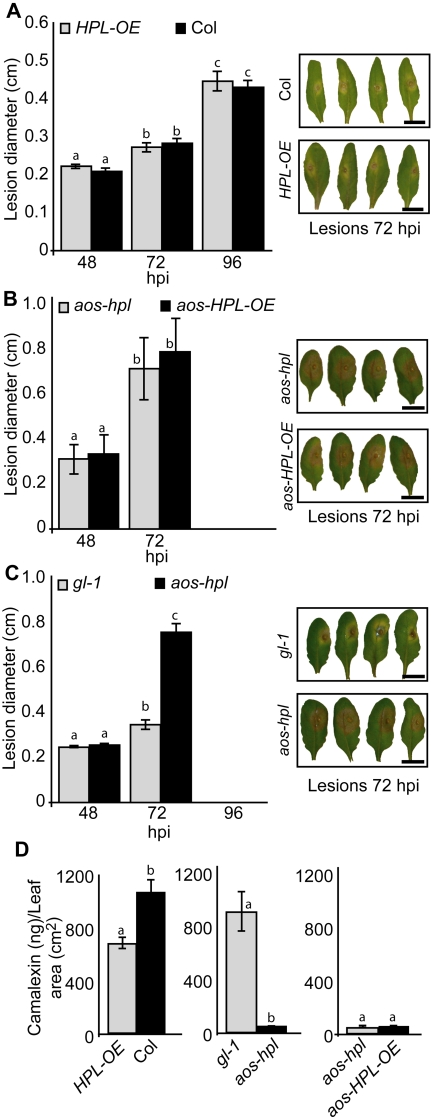
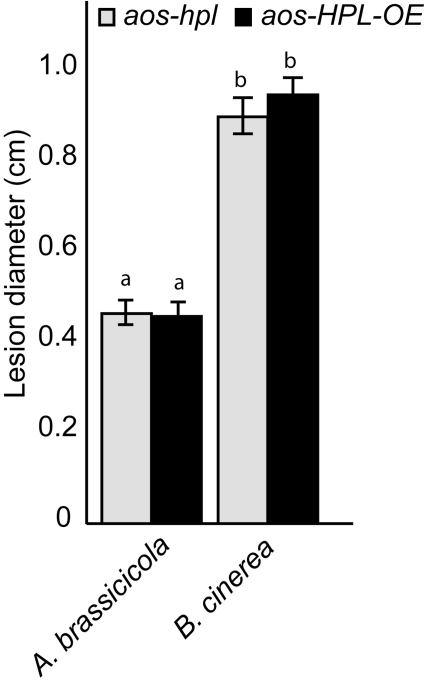
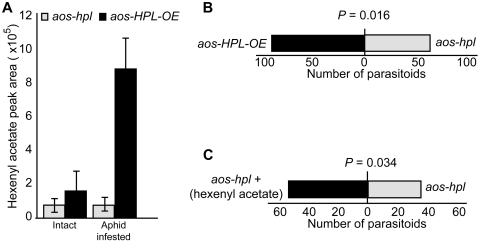
Background: Many inducible plant-defense responses are activated by jasmonates (JAs), C(6)-aldehydes, and their corresponding derivatives, produced by the two main competing branches of the oxylipin pathway, the allene oxide synthase (AOS) and hydroperoxide lyase (HPL) branches, respectively. In addition to competition for substrates, these branch-pathway-derived metabolites have substantial overlap in regulation of gene expression. Past experiments to define the role of C(6)-aldehydes in plant defense responses were biased towards the exogenous application of the synthetic metabolites or the use of genetic manipulation of HPL expression levels in plant genotypes with intact ability to produce the competing AOS-derived metabolites. To uncouple the roles of the C(6)-aldehydes and jasmonates in mediating direct and indirect plant-defense responses, we generated Arabidopsis genotypes lacking either one or both of these metabolites. These genotypes were subsequently challenged with a phloem-feeding insect (aphids: Myzus persicae), an insect herbivore (leafminers: Liriomyza trifolii), and two different necrotrophic fungal pathogens (Botrytis cinerea and Alternaria brassicicola). We also characterized the volatiles emitted by these plants upon aphid infestation or mechanical wounding and identified hexenyl acetate as the predominant compound in these volatile blends. Subsequently, we examined the signaling role of this compound in attracting the parasitoid wasp (Aphidius colemani), a natural enemy of aphids.
Principal findings: This study conclusively establishes that jasmonates and C(6)-aldehydes play distinct roles in plant defense responses. The jasmonates are indispensable metabolites in mediating the activation of direct plant-defense responses, whereas the C(6)-aldehyes are not. On the other hand, hexenyl acetate, an acetylated C(6)-aldehyde, is the predominant wound-inducible volatile signal that mediates indirect defense responses by directing tritrophic (plant-herbivore-natural enemy) interactions.
Significance: The data suggest that jasmonates and hexenyl acetate play distinct roles in mediating direct and indirect plant-defense responses. The potential advantage of this "division of labor" is to ensure the most effective defense strategy that minimizes incurred damages at a reduced metabolic cost.
Conflict of interest statement
Figures
Similar articles
-
Insect herbivores selectively suppress the HPL branch of the oxylipin pathway in host plants.Plant J. 2013 Feb;73(4):653-62. doi: 10.1111/tpj.12064. Epub 2012 Dec 28. Plant J. 2013. PMID: 23134585
-
The hydroperoxide lyase branch of the oxylipin pathway protects against photoinhibition of photosynthesis.Planta. 2017 Jun;245(6):1179-1192. doi: 10.1007/s00425-017-2674-z. Epub 2017 Mar 16. Planta. 2017. PMID: 28303390
-
Activation of the jasmonic acid pathway by depletion of the hydroperoxide lyase OsHPL3 reveals crosstalk between the HPL and AOS branches of the oxylipin pathway in rice.PLoS One. 2012;7(11):e50089. doi: 10.1371/journal.pone.0050089. Epub 2012 Nov 29. PLoS One. 2012. PMID: 23209649 Free PMC article.
-
ALLENE OXIDE SYNTHASE and HYDROPEROXIDE LYASE, Two Non-Canonical Cytochrome P450s in Arabidopsis thaliana and Their Different Roles in Plant Defense.Int J Mol Sci. 2019 Jun 23;20(12):3064. doi: 10.3390/ijms20123064. Int J Mol Sci. 2019. PMID: 31234561 Free PMC article. Review.
-
Jasmonates induce both defense responses and communication in monocotyledonous and dicotyledonous plants.Plant Cell Physiol. 2015 Jan;56(1):16-27. doi: 10.1093/pcp/pcu158. Epub 2014 Nov 4. Plant Cell Physiol. 2015. PMID: 25378688 Review.
Cited by
-
The Ectopic Expression of CaRop1 Modulates the Response of Tobacco Plants to Ralstonia solanacearum and Aphids.Front Plant Sci. 2016 Aug 8;7:1177. doi: 10.3389/fpls.2016.01177. eCollection 2016. Front Plant Sci. 2016. PMID: 27551287 Free PMC article.
-
Lipid profiling of the Arabidopsis hypersensitive response reveals specific lipid peroxidation and fragmentation processes: biogenesis of pimelic and azelaic acid.Plant Physiol. 2012 Sep;160(1):365-78. doi: 10.1104/pp.112.202846. Epub 2012 Jul 22. Plant Physiol. 2012. PMID: 22822212 Free PMC article.
-
Diversity of the enzymatic activity in the lipoxygenase gene family of Arabidopsis thaliana.Lipids. 2009 Feb;44(2):85-95. doi: 10.1007/s11745-008-3245-7. Epub 2008 Oct 24. Lipids. 2009. PMID: 18949503
-
Comparative analysis of salicylic acid levels and gene expression in resistant, tolerant, and susceptible cassava varieties following whitefly-mediated SLCMV infection.Sci Rep. 2023 Aug 21;13(1):13610. doi: 10.1038/s41598-023-40874-3. Sci Rep. 2023. PMID: 37604906 Free PMC article.
-
Cajanus cajan shows multiple novel adaptations in response to regular mechanical stress.J Plant Res. 2022 Nov;135(6):809-821. doi: 10.1007/s10265-022-01414-8. Epub 2022 Oct 15. J Plant Res. 2022. PMID: 36241771
References
-
- Paiva NL. An Introduction to the Biosynthesis of Chemicals Used in Plant-Microbe Communication. J Plant Growth Regul. 2000;19:131–143. - PubMed
-
- Walling LL. The Myriad Plant Responses to Herbivores. J Plant Growth Regul. 2000;19:195–216. - PubMed
-
- Schoonhoven LM, Jermy T, Van Loon JJA. Insect-Plant Biology– from Physiology to Evolution. Plant Growth Regulation. 1998;28:217–218.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
