

MED1 phosphorylation promotes its association with mediator: implications for nuclear receptor signaling
- PMID: 18391015
- PMCID: PMC2423130
- DOI: 10.1128/MCB.02191-07
MED1 phosphorylation promotes its association with mediator: implications for nuclear receptor signaling
Abstract
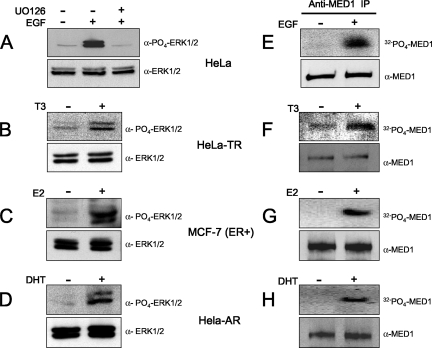
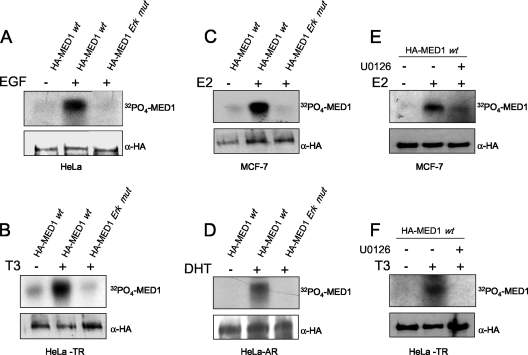
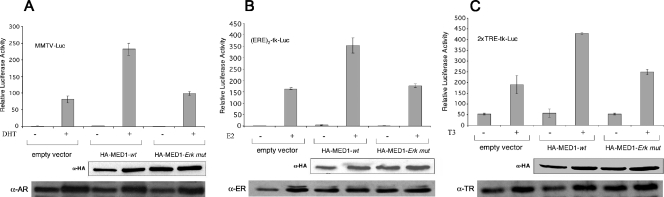
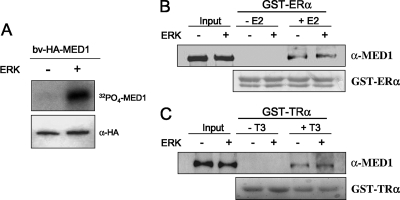
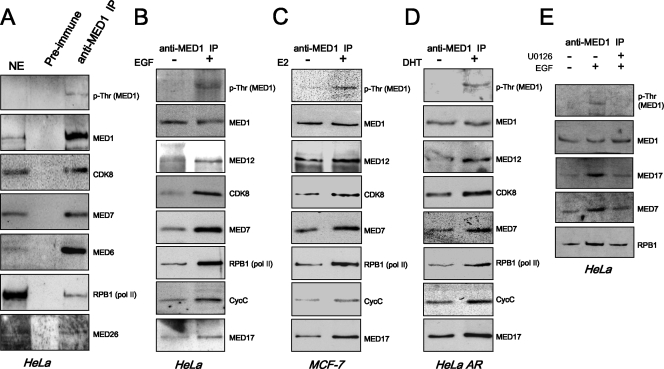
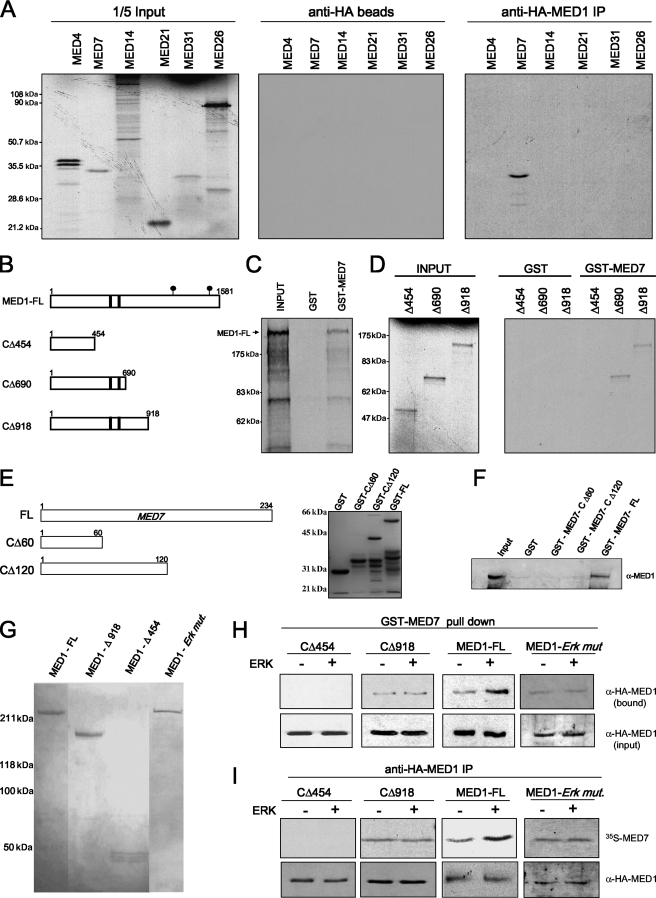
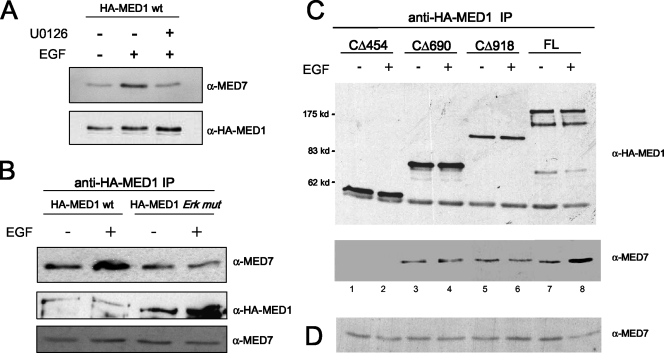
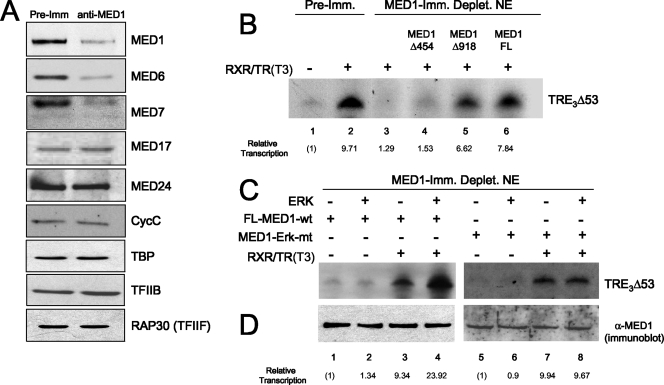
Mediator is a conserved multisubunit complex that acts as a functional interface between regulatory transcription factors and the general RNA polymerase II initiation apparatus. MED1 is a pivotal component of the complex that binds to nuclear receptors and a broad array of other gene-specific activators. Paradoxically, MED1 is found in only a fraction of the total cellular Mediator complexes, and the mechanisms regulating its binding to the core complex remain unclear. Here, we report that phosphorylation of MED1 by mitogen-activated protein kinase-extracellular signal-regulated kinase (MAPK-ERK) promotes its association with Mediator. We show that MED1 directly binds to the MED7 subunit and that ERK phosphorylation of MED1 enhances this interaction. Interestingly, we found that both thyroid and steroid hormones stimulate MED1 phosphorylation in vivo and that MED1 phosphorylation is required for its nuclear hormone receptor coactivator activity. Finally, we show that MED1 phosphorylation by ERK enhances thyroid hormone receptor-dependent transcription in vitro. Our findings suggest that ERK phosphorylation of MED1 is a regulatory mechanism that promotes MED1 association with Mediator and, as such, may facilitate a novel feed-forward action of nuclear hormones.
Figures
References
-
- Belakavadi, M., and J. D. Fondell. 2006. Role of the mediator complex in nuclear hormone receptor signaling. Rev. Physiol. Biochem. Pharmacol. 15623-43. - PubMed
-
- Chadick, J. Z., and F. J. Asturias. 2005. Structure of eukaryotic Mediator complexes. Trends Biochem. Sci. 30264-271. - PubMed
-
- Cheskis, B. J. 2004. Regulation of cell signalling cascades by steroid hormones. J. Cell Biochem. 9320-27. - PubMed
-
- Conaway, J. W., L. Florens, S. Sato, C. Tomomori-Sato, T. J. Parmely, T. Yao, S. K. Swanson, C. A. Banks, M. P. Washburn, and R. C. Conaway. 2005. The mammalian Mediator complex. FEBS Lett. 579904-908. - PubMed
-
- Crawford, S. E., C. Qi, P. Misra, V. Stellmach, M. S. Rao, J. D. Engel, Y. Zhu, and J. K. Reddy. 2002. Defects of the heart, eye, and megakaryocytes in peroxisome proliferator activator receptor-binding protein (PBP) null embryos implicate GATA family of transcription factors. J. Biol. Chem. 2773585-3592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous