

Cyclic AMP (cAMP)-mediated stimulation of adipocyte differentiation requires the synergistic action of Epac- and cAMP-dependent protein kinase-dependent processes
- PMID: 18391018
- PMCID: PMC2423297
- DOI: 10.1128/MCB.00709-07
Cyclic AMP (cAMP)-mediated stimulation of adipocyte differentiation requires the synergistic action of Epac- and cAMP-dependent protein kinase-dependent processes
Abstract
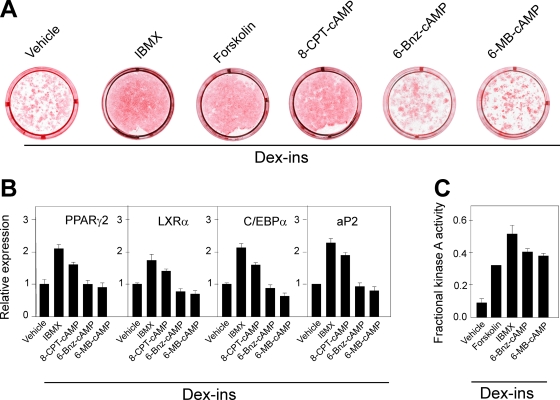
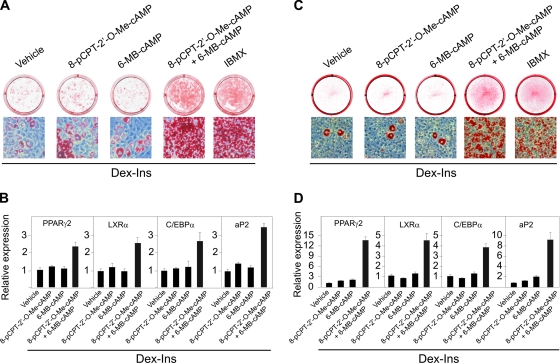
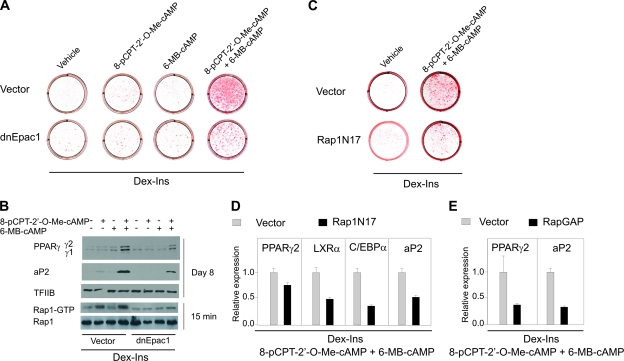
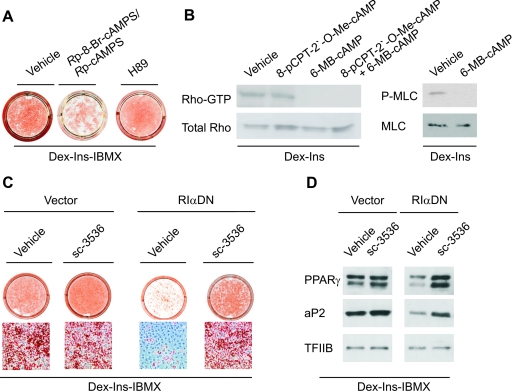
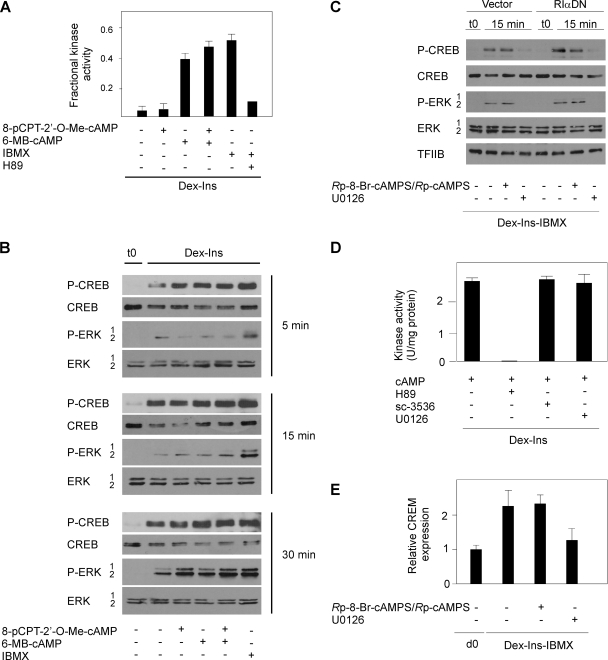
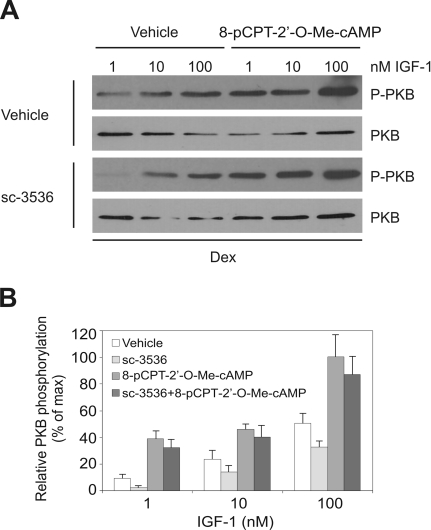
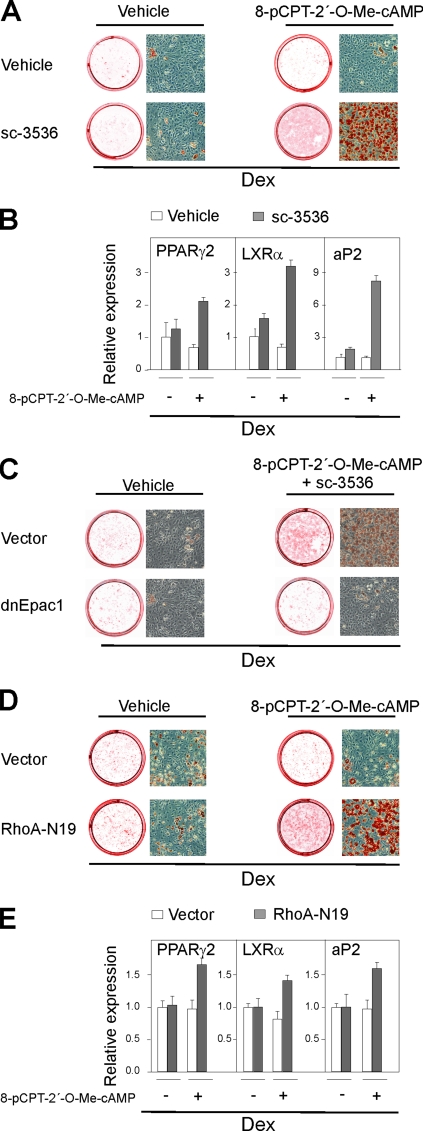
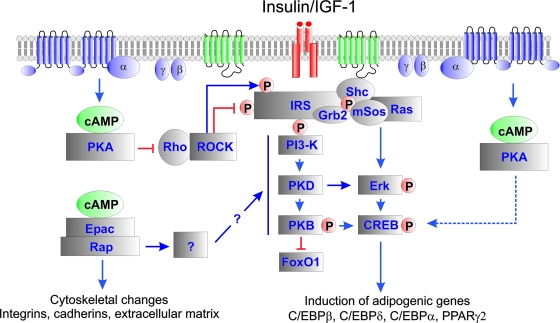
Cyclic AMP (cAMP)-dependent processes are pivotal during the early stages of adipocyte differentiation. We show that exchange protein directly activated by cAMP (Epac), which functions as a guanine nucleotide exchange factor for the Ras-like GTPases Rap1 and Rap2, was required for cAMP-dependent stimulation of adipocyte differentiation. Epac, working via Rap, acted synergistically with cAMP-dependent protein kinase (protein kinase A [PKA]) to promote adipogenesis. The major role of PKA was to down-regulate Rho and Rho-kinase activity, rather than to enhance CREB phosphorylation. Suppression of Rho-kinase impaired proadipogenic insulin/insulin-like growth factor 1 signaling, which was restored by activation of Epac. This interplay between PKA and Epac-mediated processes not only provides novel insight into the initiation and tuning of adipocyte differentiation, but also demonstrates a new mechanism of cAMP signaling whereby cAMP uses both PKA and Epac to achieve an appropriate cellular response.
Figures

Similar articles
-
Cyclic AMP induces integrin-mediated cell adhesion through Epac and Rap1 upon stimulation of the beta 2-adrenergic receptor.J Cell Biol. 2003 Feb 17;160(4):487-93. doi: 10.1083/jcb.200209105. Epub 2003 Feb 10. J Cell Biol. 2003. PMID: 12578910 Free PMC article.
-
Epac, in synergy with cAMP-dependent protein kinase (PKA), is required for cAMP-mediated mitogenesis.J Biol Chem. 2008 Feb 22;283(8):4464-8. doi: 10.1074/jbc.C700171200. Epub 2007 Dec 6. J Biol Chem. 2008. PMID: 18063584
-
Activation of G protein-coupled estrogen receptor 1 induces coronary artery relaxation via Epac/Rap1-mediated inhibition of RhoA/Rho kinase pathway in parallel with PKA.PLoS One. 2017 Mar 9;12(3):e0173085. doi: 10.1371/journal.pone.0173085. eCollection 2017. PLoS One. 2017. PMID: 28278256 Free PMC article.
-
Rap-linked cAMP signaling Epac proteins: compartmentation, functioning and disease implications.Cell Signal. 2011 Aug;23(8):1257-66. doi: 10.1016/j.cellsig.2011.03.007. Epub 2011 Mar 22. Cell Signal. 2011. PMID: 21402149 Review.
-
Insights into exchange factor directly activated by cAMP (EPAC) as potential target for cancer treatment.Mol Cell Biochem. 2018 Oct;447(1-2):77-92. doi: 10.1007/s11010-018-3294-z. Epub 2018 Feb 7. Mol Cell Biochem. 2018. PMID: 29417338 Review.
Cited by
-
Chromatin accessibility and transcription factor binding at the PPARγ2 promoter during adipogenesis is protein kinase A-dependent.J Cell Physiol. 2011 Jan;226(1):86-93. doi: 10.1002/jcp.22308. J Cell Physiol. 2011. PMID: 20625991 Free PMC article.
-
Distinct signalling properties of insulin receptor substrate (IRS)-1 and IRS-2 in mediating insulin/IGF-1 action.Cell Signal. 2018 Jul;47:1-15. doi: 10.1016/j.cellsig.2018.03.003. Epub 2018 Mar 14. Cell Signal. 2018. PMID: 29550500 Free PMC article.
-
Dynamic mitochondrial-nuclear redistribution of the immunophilin FKBP51 is regulated by the PKA signaling pathway to control gene expression during adipocyte differentiation.J Cell Sci. 2013 Dec 1;126(Pt 23):5357-68. doi: 10.1242/jcs.125799. Epub 2013 Oct 7. J Cell Sci. 2013. PMID: 24101724 Free PMC article.
-
Epac, not PKA catalytic subunit, is required for 3T3-L1 preadipocyte differentiation.Front Biosci (Elite Ed). 2010 Jan 1;2(2):392-8. doi: 10.2741/e99. Front Biosci (Elite Ed). 2010. PMID: 20036887 Free PMC article.
-
Protein-coding variants implicate novel genes related to lipid homeostasis contributing to body-fat distribution.Nat Genet. 2019 Mar;51(3):452-469. doi: 10.1038/s41588-018-0334-2. Epub 2019 Feb 18. Nat Genet. 2019. PMID: 30778226 Free PMC article.
References
-
- Baisamy, L., N. Jurisch, and D. Diviani. 2005. Leucine zipper-mediated homo-oligomerization regulates the Rho-GEF activity of AKAP-Lbc. J. Biol. Chem. 28015405-15412. - PubMed
-
- Begum, N., O. A. Sandu, and N. Duddy. 2002. Negative regulation of rho signaling by insulin and its impact on actin on cytoskeleton organization in vascular smooth muscle cells: role of nitric oxide and cyclic guanosine monophosphate signaling pathways. Diabetes 512256-2263. - PubMed
-
- Begum, N., O. A. Sandu, M. Ito, S. M. Lohmann, and A. Smolenski. 2002. Active Rho kinase (ROK-alpha) associates with insulin receptor substrate-1 and inhibits insulin signaling in vascular smooth muscle cells. J. Biol. Chem. 2776214-6222. - PubMed
-
- Belmonte, N., B. W. Phillips, F. Massiera, P. Villageois, B. Wdziekonski, P. Saint-Marc, J. Nichols, J. Aubert, K. Saeki, A. Yuo, S. Narumiya, G. Ailhaud, and C. Dani. 2001. Activation of extracellular signal-regulated kinases and CREB/ATF-1 mediate the expression of CCAAT/enhancer binding proteins β and -δ in preadipocytes. Mol. Endocrinol. 152037-2049. - PubMed
-
- Bennett, C. N., S. E. Ross, K. A. Longo, L. Bajnok, N. Hemati, K. W. Johnson, S. D. Harrison, and O. A. MacDougald. 2002. Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 27730998-31004. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources