

Compensatory role for Pyk2 during angiogenesis in adult mice lacking endothelial cell FAK
- PMID: 18391070
- PMCID: PMC2287283
- DOI: 10.1083/jcb.200710038
Compensatory role for Pyk2 during angiogenesis in adult mice lacking endothelial cell FAK
Abstract
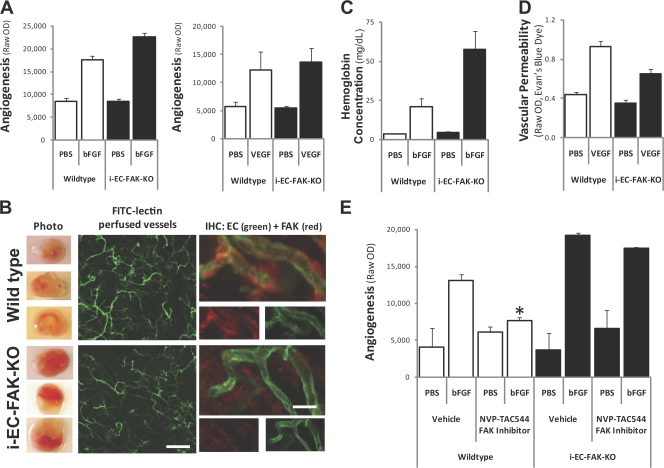
Focal adhesion kinase (FAK) plays a critical role during vascular development because knockout of FAK in endothelial cells (ECs) is embryonic lethal. Surprisingly, tamoxifen-inducible conditional knockout of FAK in adult blood vessels (inducible EC-specific FAK knockout [i-EC-FAK-KO]) produces no vascular phenotype, and these animals are capable of developing a robust growth factor-induced angiogenic response. Although angiogenesis in wild-type mice is suppressed by pharmacological inhibition of FAK, i-EC-FAK-KO mice are refractory to this treatment, which suggests that adult i-EC-FAK-KO mice develop a compensatory mechanism to bypass the requirement for FAK. Indeed, expression of the FAK-related proline-rich tyrosine kinase 2 (Pyk2) is elevated and phosphorylated in i-EC-FAK-KO blood vessels. In cultured ECs, FAK knockdown leads to increased Pyk2 expression and, surprisingly, FAK kinase inhibition leads to increased Pyk2 phosphorylation. Pyk2 can functionally compensate for the loss of FAK because knockdown or pharmacological inhibition of Pyk2 disrupts angiogenesis in i-EC-FAK-KO mice. These studies reveal the adaptive capacity of ECs to switch to Pyk2-dependent signaling after deletion or kinase inhibition of FAK.
Figures




References
-
- Allingham, M.J., J.D. van Buul, and K. Burridge. 2007. ICAM-1-mediated, Src- and Pyk2-dependent vascular endothelial cadherin tyrosine phosphorylation is required for leukocyte transendothelial migration. J. Immunol. 179:4053–4064. - PubMed
-
- Avraham, H.K., T.H. Lee, Y. Koh, T.A. Kim, S. Jiang, M. Sussman, A.M. Samarel, and S. Avraham. 2003. Vascular endothelial growth factor regulates focal adhesion assembly in human brain microvascular endothelial cells through activation of the focal adhesion kinase and related adhesion focal tyrosine kinase. J. Biol. Chem. 278:36661–36668. - PubMed
-
- Brooks, P.C., R.A. Clark, and D.A. Cheresh. 1994. Requirement of vascular integrin alpha v beta 3 for angiogenesis. Science. 264:569–571. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
