

TFIID component TAF7 functionally interacts with both TFIIH and P-TEFb
- PMID: 18391197
- PMCID: PMC2291086
- DOI: 10.1073/pnas.0801637105
TFIID component TAF7 functionally interacts with both TFIIH and P-TEFb
Abstract
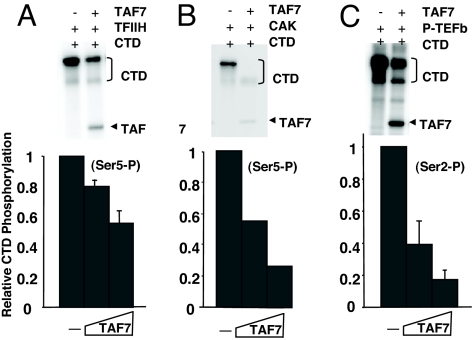
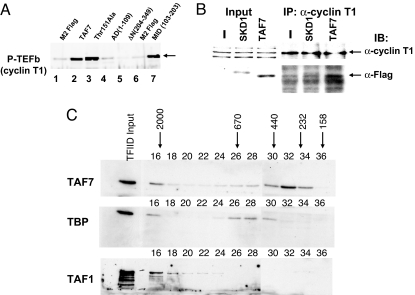
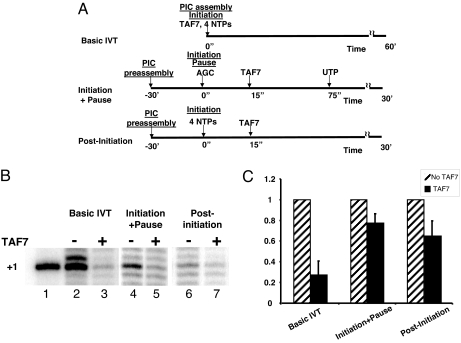
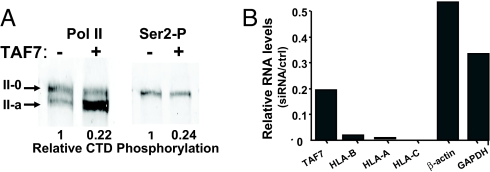
Transcription consists of a series of highly regulated steps: assembly of the preinitiation complex (PIC) at the promoter, initiation, elongation, and termination. PIC assembly is nucleated by TFIID, a complex composed of the TATA-binding protein (TBP) and a series of TBP-associated factors (TAFs). One component, TAF7, is incorporated in the PIC through its interaction with TFIID but is released from TFIID upon transcription initiation. We now report that TAF7 interacts with the transcription factors, TFIIH and P-TEFb, resulting in the inhibition of their Pol II CTD kinase activities. Importantly, in in vitro transcription reactions, TAF7 inhibits steps after PIC assembly and formation of the first phosphodiester bonds. Further, in vivo TAF7 coelongates with P-TEFb and Pol II downstream of the promoter. We propose a model in which TAF7 contributes to the regulation of the transition from PIC assembly to initiation and elongation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Woychik NA, Hampsey M. The RNA polymerase II machinery: Structure illuminates function. Cell. 2002;108:453–463. - PubMed
-
- Zawel L, Kumar KP, Reinberg D. Recycling of the general transcription factors during RNA polymerase II transcription. Genes Dev. 1995;9:1479–1490. - PubMed
-
- Yudkovsky N, Ranish JA, Hahn S. A transcription reinitiation intermediate that is stabilized by activator. Nature. 2000;408:225–229. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
