

Structure of human monoamine oxidase A at 2.2-A resolution: the control of opening the entry for substrates/inhibitors
- PMID: 18391214
- PMCID: PMC2311356
- DOI: 10.1073/pnas.0710626105
Structure of human monoamine oxidase A at 2.2-A resolution: the control of opening the entry for substrates/inhibitors
Abstract
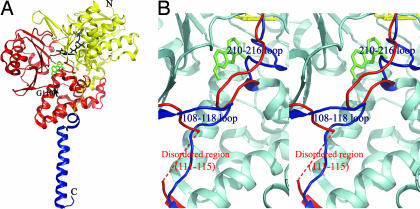
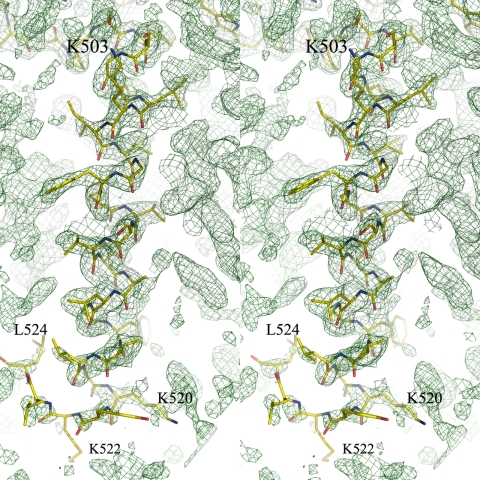
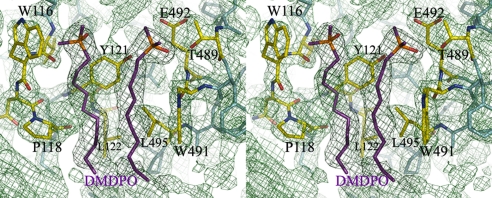
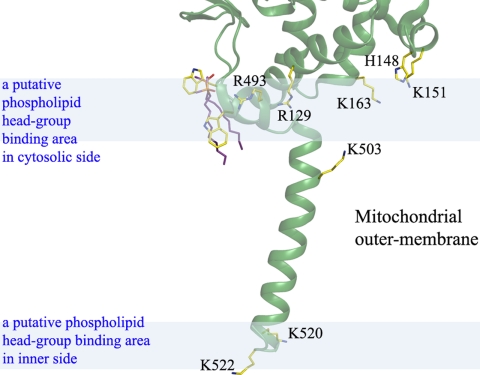
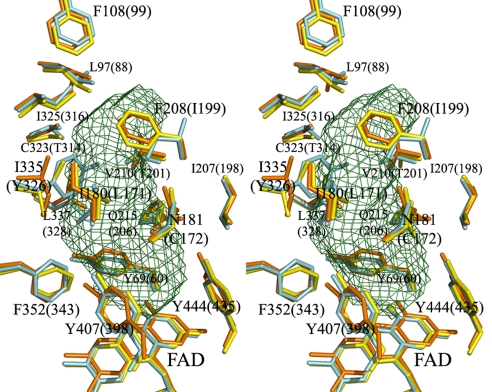
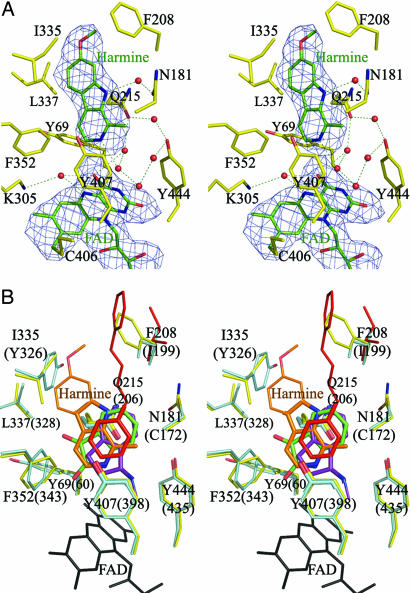
The mitochondrial outer membrane-anchored monoamine oxidase (MAO) is a biochemically important flavoenzyme that catalyzes the deamination of biogenic and xenobiotic amines. Its two subtypes, MAOA and MAOB, are linked to several psychiatric disorders and therefore are interesting targets for drug design. To understand the relationship between structure and function of this enzyme, we extended our previous low-resolution rat MAOA structure to the high-resolution wild-type and G110A mutant human MAOA structures at 2.2 and 2.17 A, respectively. The high-resolution MAOA structures are similar to those of rat MAOA and human MAOB, but different from the known structure of human MAOA [De Colibus L, et al. (2005) Proc Natl Acad Sci USA 102:12684-12689], specifically regarding residues 108-118 and 210-216, which surround the substrate/inhibitor cavity. The results confirm that the inhibitor selectivity of MAOA and MAOB is caused by the structural differences arising from Ile-335 in MAOA vs. Tyr-326 in MAOB. The structures exhibit a C-terminal transmembrane helix with clear electron density, as is also seen in rat MAOA. Mutations on one residue of loop 108-118, G110, which is far from the active center but close to the membrane surface, cause the solubilized enzyme to undergo a dramatic drop in activity, but have less effect when the enzyme is anchored in the membrane. These results suggest that the flexibility of loop 108-118, facilitated by anchoring the enzyme into the membrane, is essential for controlling substrate access to the active site. We report on the observation of the structure-function relationship between a transmembrane helical anchor and an extra-membrane domain.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Walker WH, Kearney EB, Seng RL, Singer TP. The covalently bound flavin of hepatic monoamine oxidase. 2. Identification and properties of cysteinyl riboflavin. Eur J Biochem. 1971;24:328–331. - PubMed
-
- Youdim MBH, Riederer P. The relevance of glial monoamine oxidase B and polyamines to the action of selegiline in Parkinson's disease. Movement Disorders. 1993;8:S8–S13. - PubMed
-
- Saura J, et al. Localization of monoamine oxidases in human peripheral tissues. Life Sci. 1996;59:1341–1349. - PubMed
-
- Binda C, Newton-Vinson P, Hubalek F, Edmondson DE, Mattevi A. Structure of human monoamine oxidase B, a drug target for the treatment of neurological disorders. Nat Struct Biol. 2002;9:22–26. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
