

S-adenosylmethionine inhibits lipopolysaccharide-induced gene expression via modulation of histone methylation
- PMID: 18393372
- PMCID: PMC2408693
- DOI: 10.1002/hep.22231
S-adenosylmethionine inhibits lipopolysaccharide-induced gene expression via modulation of histone methylation
Abstract
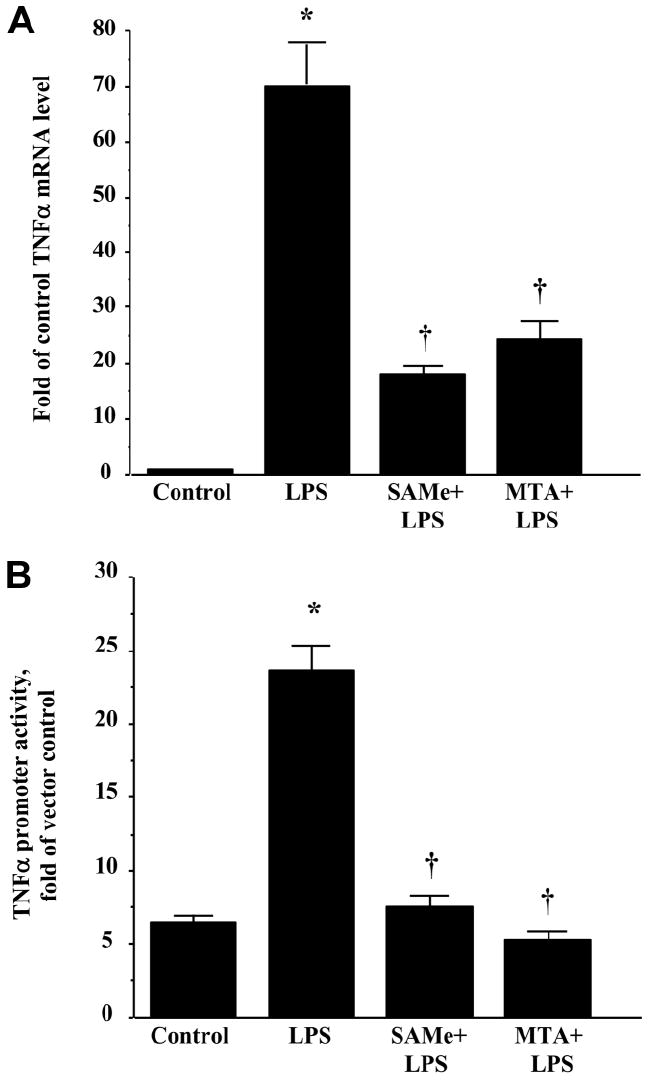
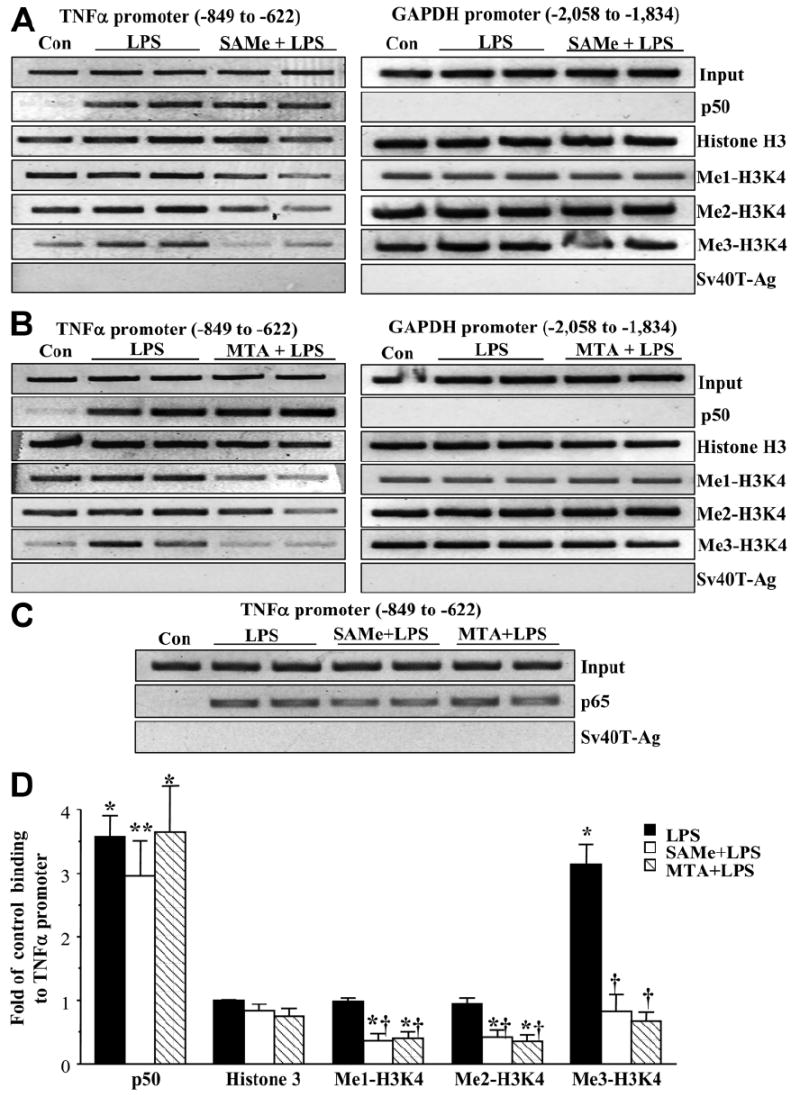
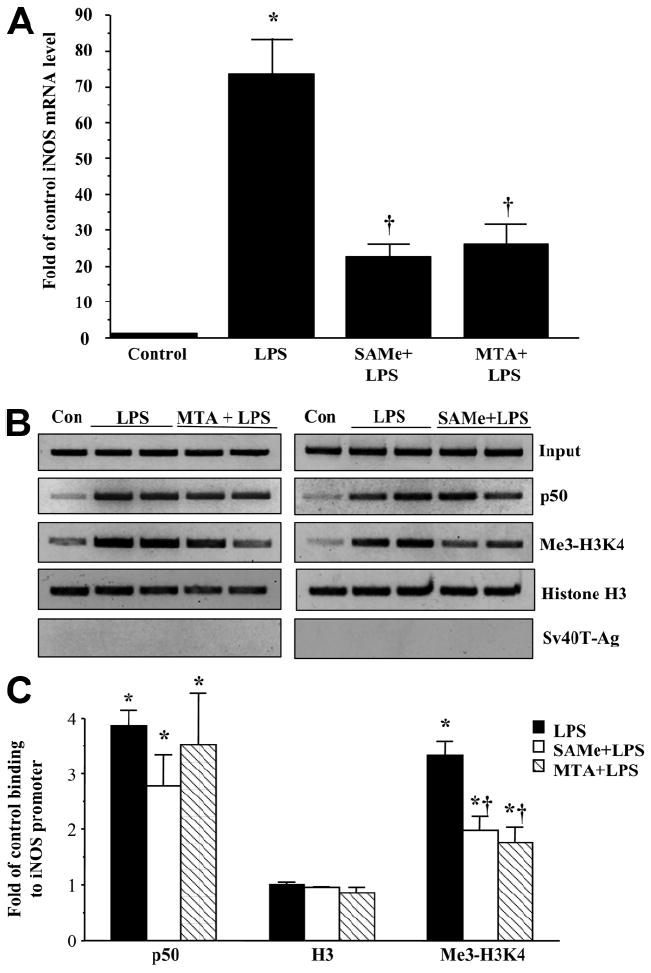
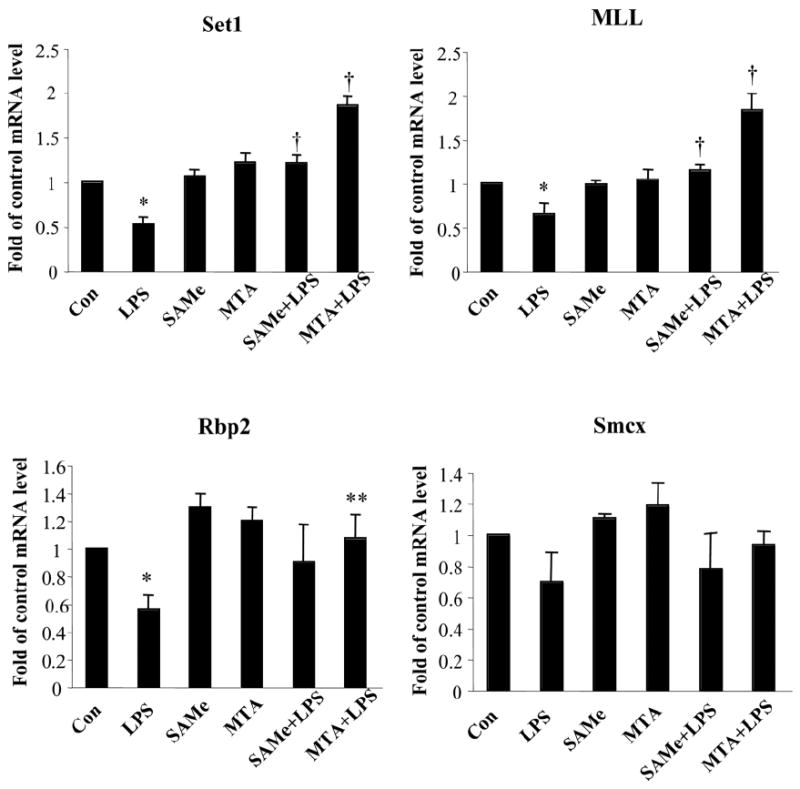
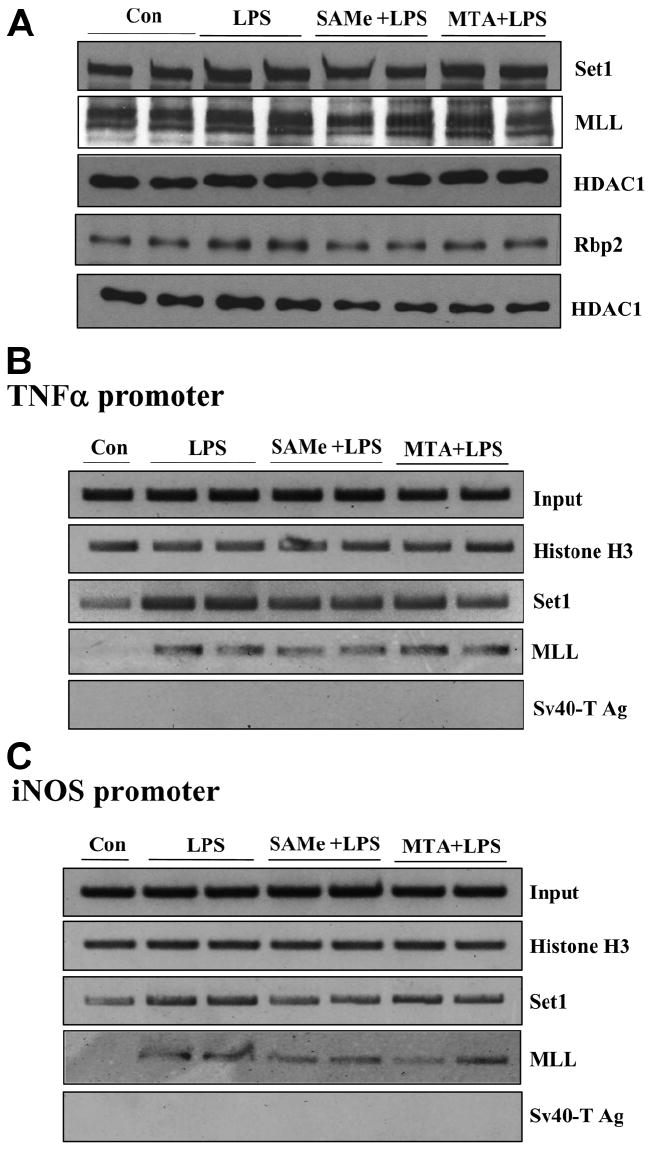
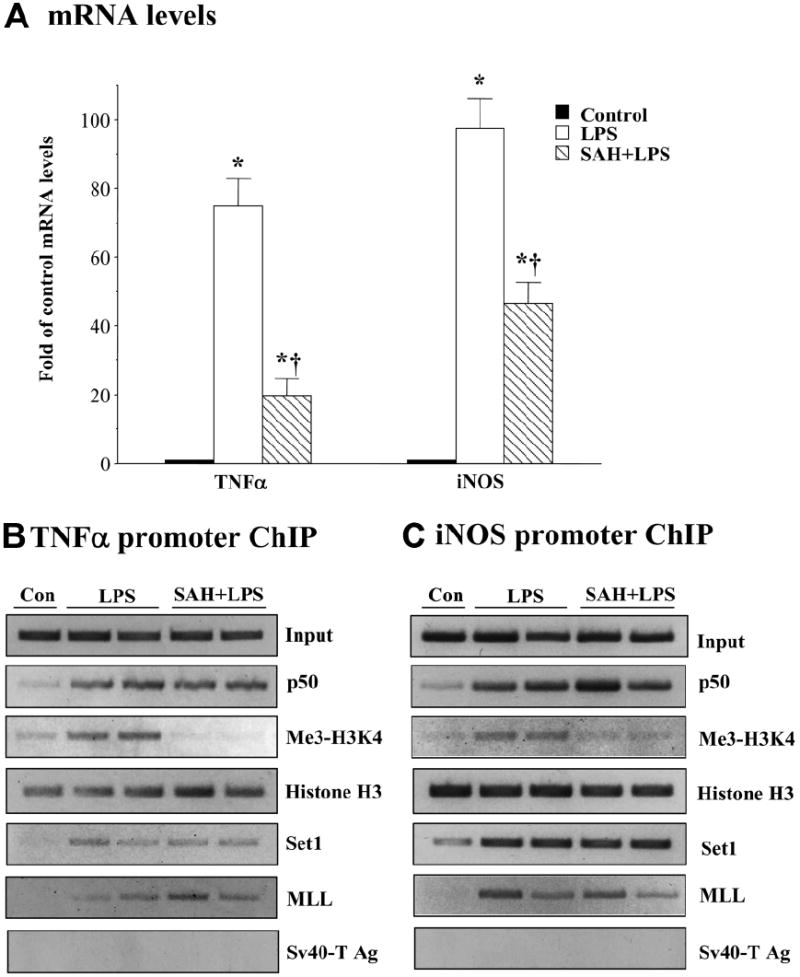
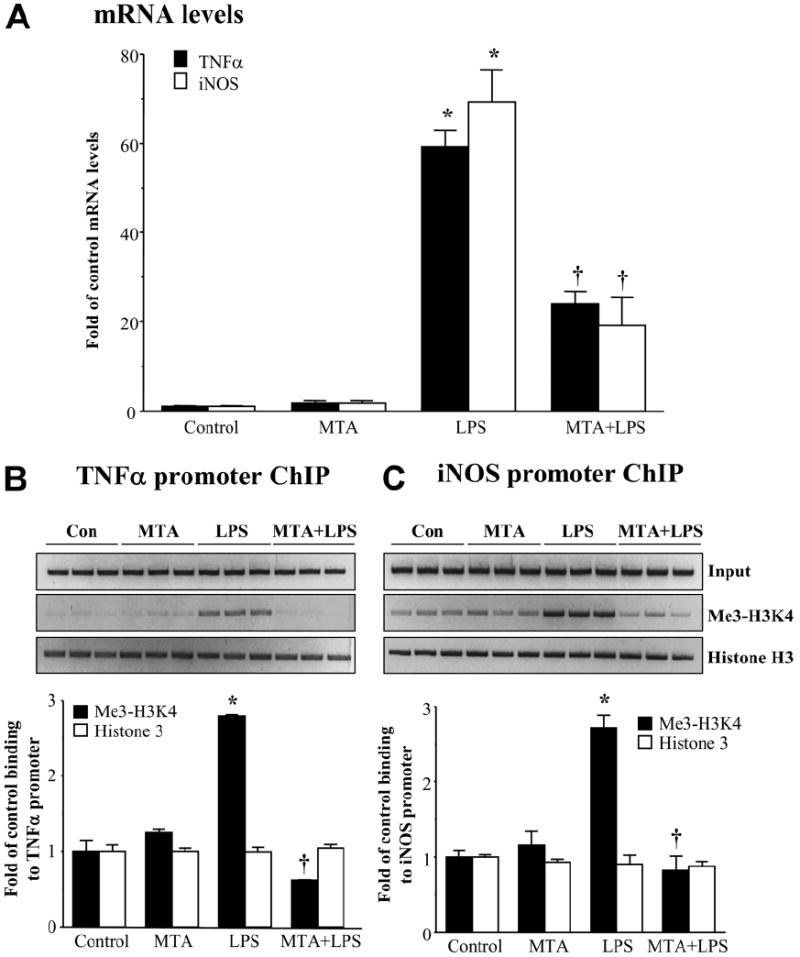
We previously showed that S-adenosylmethionine (SAMe) and its metabolite methylthioadenosine (MTA) blocked lipopolysaccharide (LPS)-induced tumor necrosis factor alpha (TNFalpha) expression in RAW (murine macrophage cell line) and Kupffer cells at the transcriptional level without affecting nuclear factor kappa B nuclear binding. However, the exact molecular mechanism or mechanisms of the inhibitory effect were unclear. While SAMe is a methyl donor, MTA is an inhibitor of methylation. SAMe can convert to MTA spontaneously, so the effect of exogenous SAMe may be mediated by MTA. The aim of our current work is to examine whether the mechanism of SAMe and MTA's inhibitory effect on proinflammatory mediators might involve modulation of histone methylation. In RAW cells, we found that LPS induced TNFalpha expression by both transcriptional and posttranscriptional mechanisms. SAMe and MTA treatment inhibited the LPS-induced increase in gene transcription. Using the chromatin immunoprecipitation assay, we found that LPS increased the binding of trimethylated histone 3 lysine 4 (H3K4) to the TNFalpha promoter, and this was completely blocked by either SAMe or MTA pretreatment. Similar effects were observed with LPS-mediated induction of inducible nitric oxide synthase (iNOS). LPS increased the binding of histone methyltransferases Set1 and myeloid/lymphoid leukemia to these promoters, which was unaffected by SAMe or MTA. The effects of MTA in RAW cells were confirmed in vivo in LPS-treated mice. Exogenous SAMe is unstable and converts spontaneously to MTA, which is stable and cell-permeant. Treatment with SAMe doubled intracellular MTA and S-adenosylhomocysteine (SAH) levels. SAH also inhibited H3K4 binding to TNFalpha and iNOS promoters.
Conclusion: The mechanism of SAMe's pharmacologic inhibitory effect on proinflammatory mediators is mainly mediated by MTA and SAH at the level of histone methylation.
Conflict of interest statement
Potential conflict of interest: Nothing to report.
Figures

References
-
- Mato JM, Lu SC. Role of S-adenosyl-L-methionine in liver health and injury. HEPATOLOGY. 2007;45:1306–1312. - PubMed
-
- Su GL. Lipopolysaccharides in liver injury: molecular mechanisms of Kupffer cell activation. Am J Physiol Gastrointest Liver Physiol. 2002;283:G256–G265. - PubMed
-
- Tracey KJ, Cerami A. Tumor necrosis factor: a pleiotropic cytokine and therapeutic target. Annu Rev Med. 1994;45:491–503. - PubMed
-
- Li J, Billiar TR. Nitric oxide. IV. Determinants of nitric oxide protection and toxicity in liver. Am J Physiol. 1999;276:G1069–G1073. - PubMed
-
- Veal N, Hsieh CL, Xiong S, Mato JM, Lu S, Tsukamoto H. Inhibition of lipopolysaccharide-stimulated TNF-alpha promoter activity by S-adenosylmethionine and 5′-methylthioadenosine. Am J Physiol Gastrointest Liver Physiol. 2004;287:G352–G362. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources