

Cell-polarity dynamics controls the mechanism of lumen formation in epithelial morphogenesis
- PMID: 18394894
- PMCID: PMC2405957
- DOI: 10.1016/j.cub.2008.02.076
Cell-polarity dynamics controls the mechanism of lumen formation in epithelial morphogenesis
Erratum in
- Curr Biol. 2008 Apr 22;18(8):630. Ewald, Andrew [corrected to Ewald, Andrew J]
- Curr Biol. 2008 Jul 8;18(13):1016
Abstract
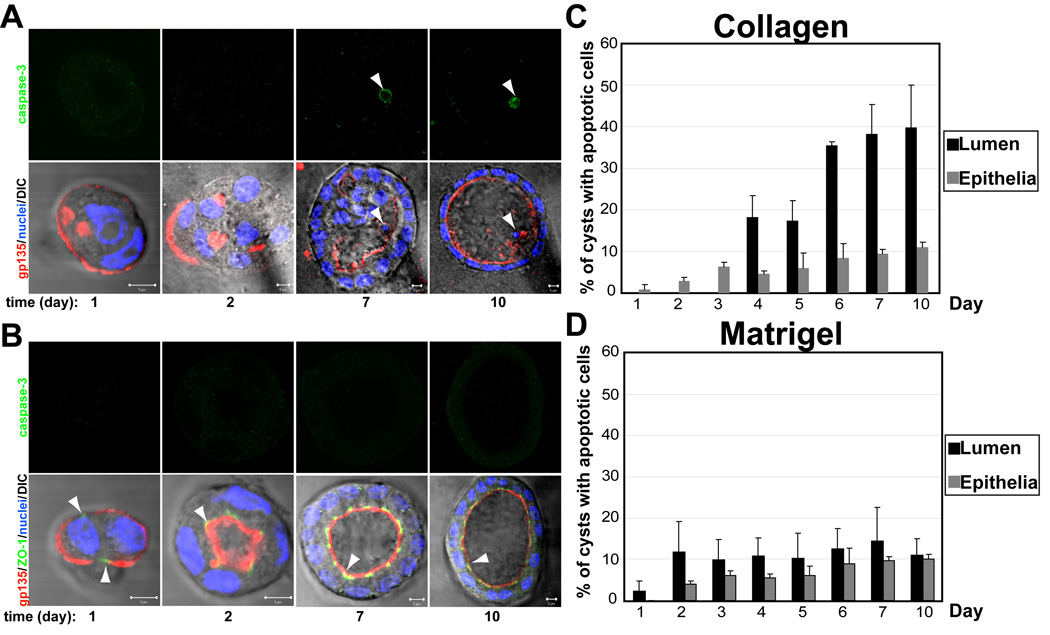
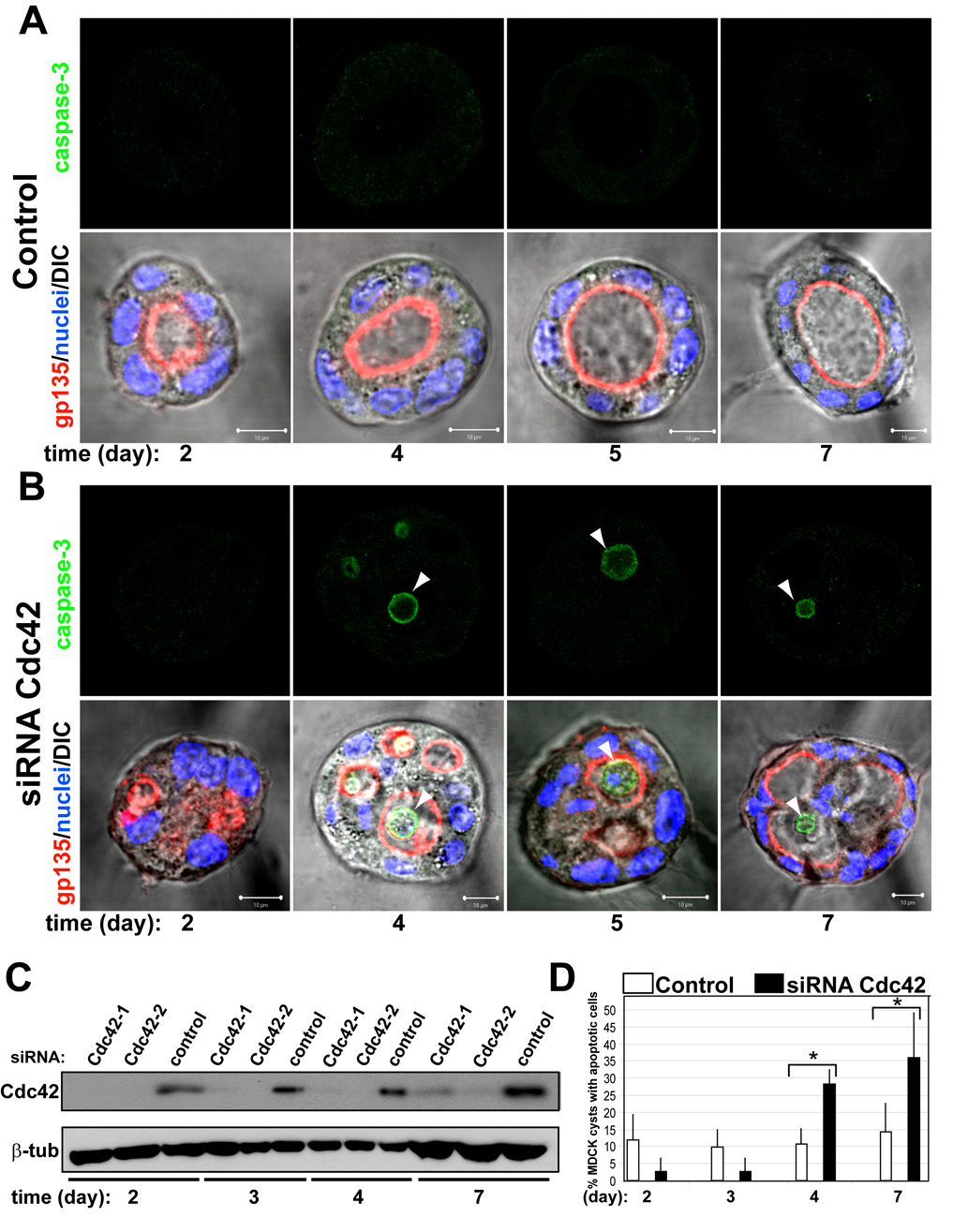
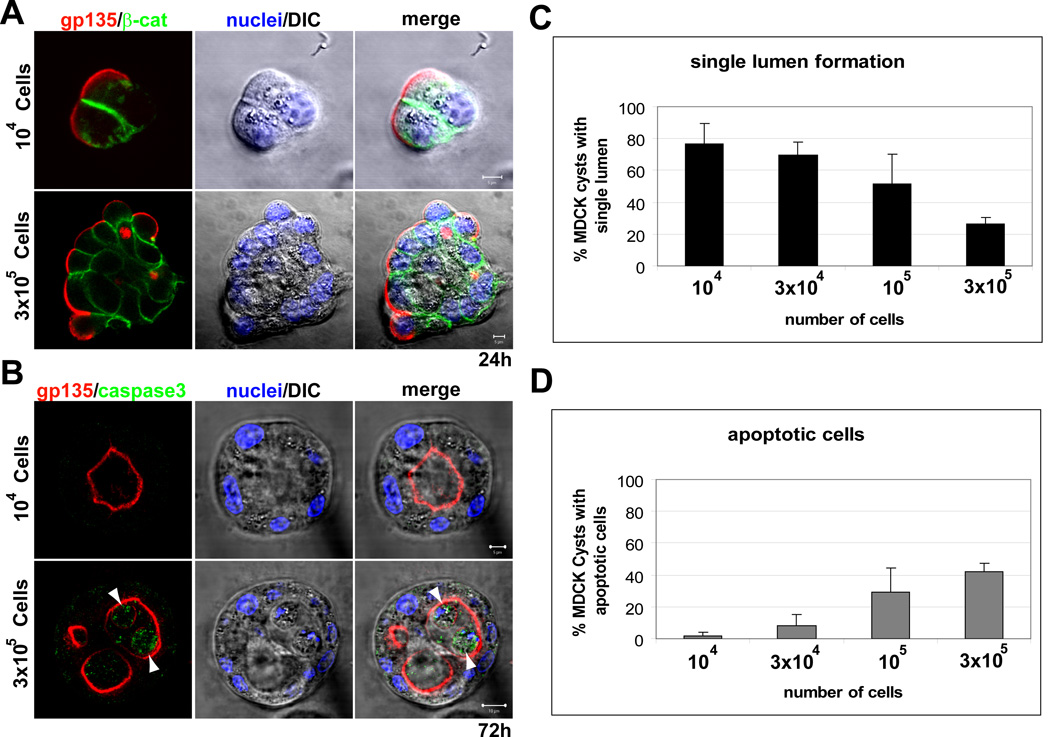
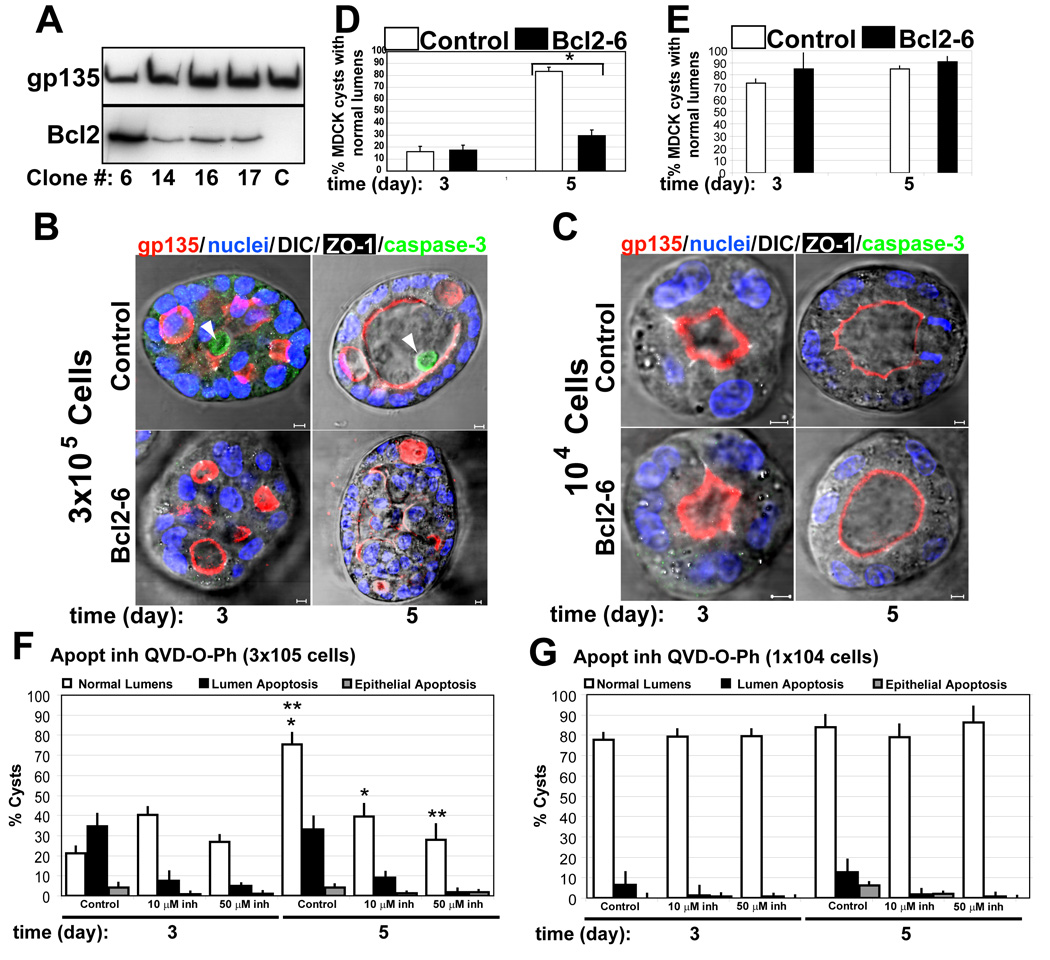
Many organs consist of tubes of epithelial cells enclosing a central lumen. How the space of this lumen is generated is a key question in morphogenesis. Two predominant mechanisms of de novo lumen formation have been observed: hollowing and cavitation. In hollowing, the lumen is formed by exocytosis and membrane separation, whereas, in cavitation, the lumen is generated by apoptosis of cells in the middle of the structure [1, 2]. Using MDCK cells in three-dimensional cultures, we found an inverse correlation between polarization efficiency and apoptosis. When cells were grown in collagen, where cells polarized slowly, apoptosis was needed for lumen formation. However, in the presence of Matrigel, which allowed rapid polarization, lumens formed without apoptosis. If polarization in Matrigel was perturbed by blocking formation of the apical surface by RNAi of Cdc42, lumens formed by apoptosis. In a complementary approach, we plated cells at high density so that aggregates formed with little polarity. These aggregates required apoptosis to form lumens, whereas cells plated at low density formed cysts with rapidly polarizing cells and did not need apoptosis to form lumens. The mechanism of lumen formation in the 3D-MDCK model can shift between hollowing and cavitation, depending on cell polarization.
Figures
References
-
- Lubarsky B, Krasnow MA. Tube morphogenesis: making and shaping biological tubes. Cell. 2003;112:19–28. - PubMed
-
- O'Brien LE, Zegers MM, Mostov KE. Opinion: Building epithelial architecture: insights from three-dimensional culture models. Nat Rev Mol Cell Biol. 2002;3:531–537. - PubMed
-
- Lin HH, Yang TP, Jiang ST, Yang HY, Tang MJ. Bcl-2 overexpression prevents apoptosis-induced Madin-Darby canine kidney simple epithelial cyst formation. Kidney Int. 1999;55:168–178. - PubMed
-
- O'Brien LE, Jou TS, Pollack AL, Zhang Q, Hansen SH, Yurchenco P, Mostov KE. Rac1 orientates epithelial apical polarity through effects on basolateral laminin assembly. Nat Cell Biol. 2001;3:831–838. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
