

Diffusion of the second messengers in the cytoplasm acts as a variability suppressor of the single photon response in vertebrate phototransduction
- PMID: 18400950
- PMCID: PMC2292384
- DOI: 10.1529/biophysj.107.114058
Diffusion of the second messengers in the cytoplasm acts as a variability suppressor of the single photon response in vertebrate phototransduction
Abstract
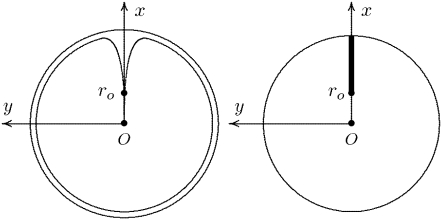
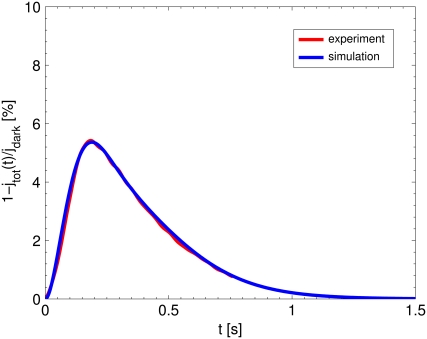
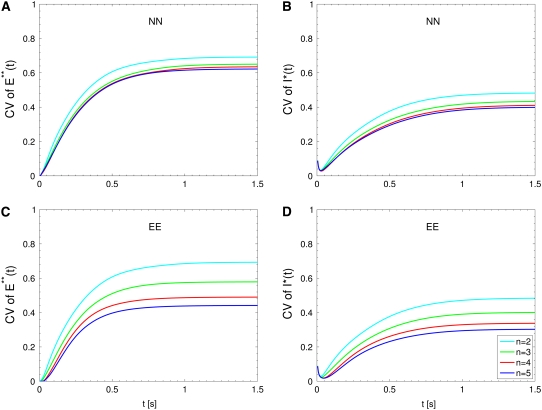
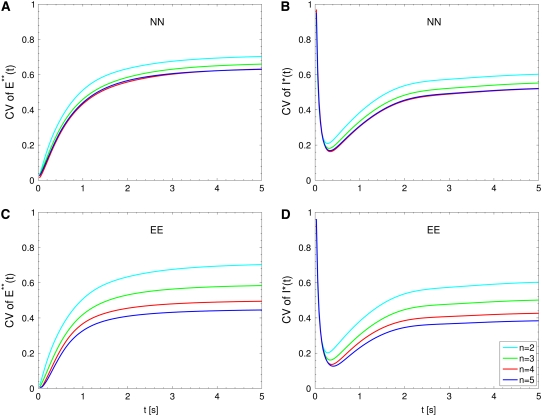
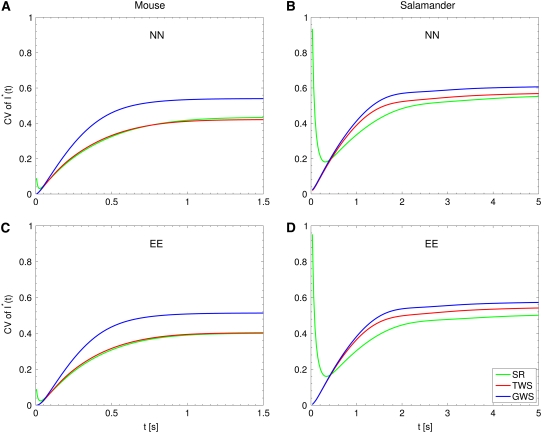
The single photon response in vertebrate phototransduction is highly reproducible despite a number of random components of the activation cascade, including the random activation site, the random walk of an activated receptor, and its quenching in a random number of steps. Here we use a previously generated and tested spatiotemporal mathematical and computational model to identify possible mechanisms of variability reduction. The model permits one to separate the process into modules, and to analyze their impact separately. We show that the activation cascade is responsible for generation of variability, whereas diffusion of the second messengers is responsible for its suppression. Randomness of the activation site contributes at early times to the coefficient of variation of the photoresponse, whereas the Brownian path of a photoisomerized rhodopsin (Rh*) has a negligible effect. The major driver of variability is the turnoff mechanism of Rh*, which occurs essentially within the first 2-4 phosphorylated states of Rh*. Theoretically increasing the number of steps to quenching does not significantly decrease the corresponding coefficient of variation of the effector, in agreement with the biochemical limitations on the phosphorylated states of the receptor. Diffusion of the second messengers in the cytosol acts as a suppressor of the variability generated by the activation cascade. Calcium feedback has a negligible regulatory effect on the photocurrent variability. A comparative variability analysis has been conducted for the phototransduction in mouse and salamander, including a study of the effects of their anatomical differences such as incisures and photoreceptors geometry on variability generation and suppression.
Figures
References
-
- Aho, A. C., K. Donner, C. Hyden, L. O. Larsen, and T. Reuter. 1988. Low retinal noise in animals with low body temperature allows high visual sensitivity. Nature. 334:348–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
