

Inflammatory tumour cell killing by oncolytic reovirus for the treatment of melanoma
- PMID: 18401435
- PMCID: PMC4821075
- DOI: 10.1038/gt.2008.58
Inflammatory tumour cell killing by oncolytic reovirus for the treatment of melanoma
Abstract
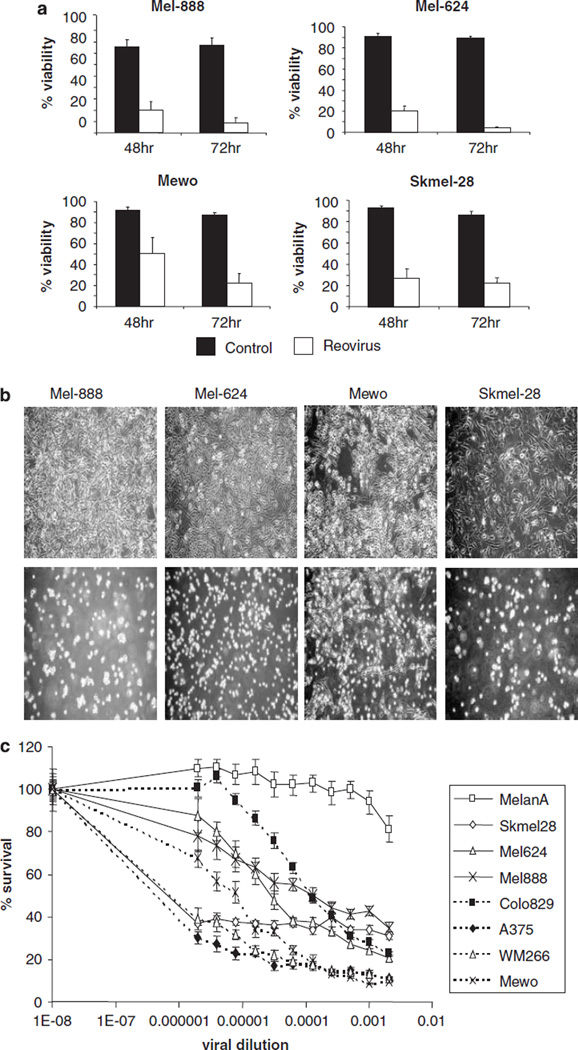
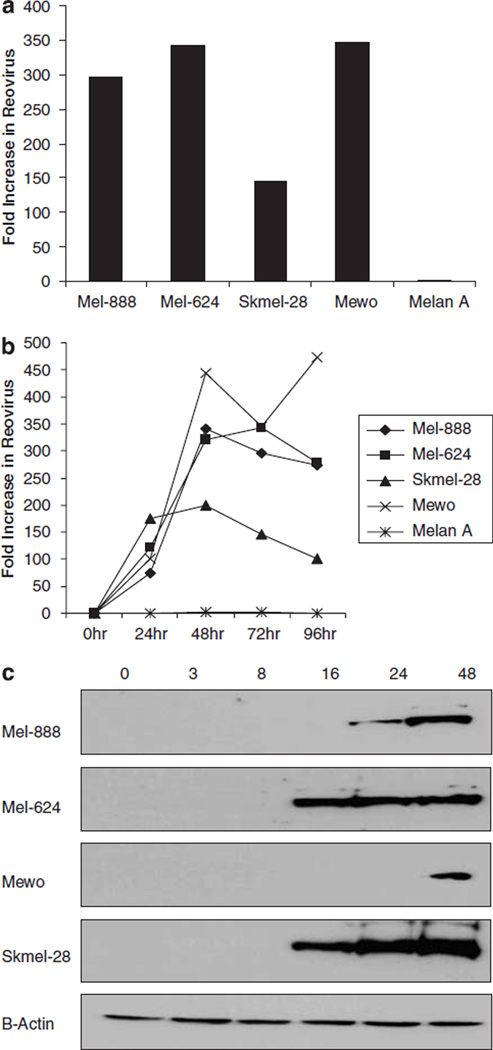
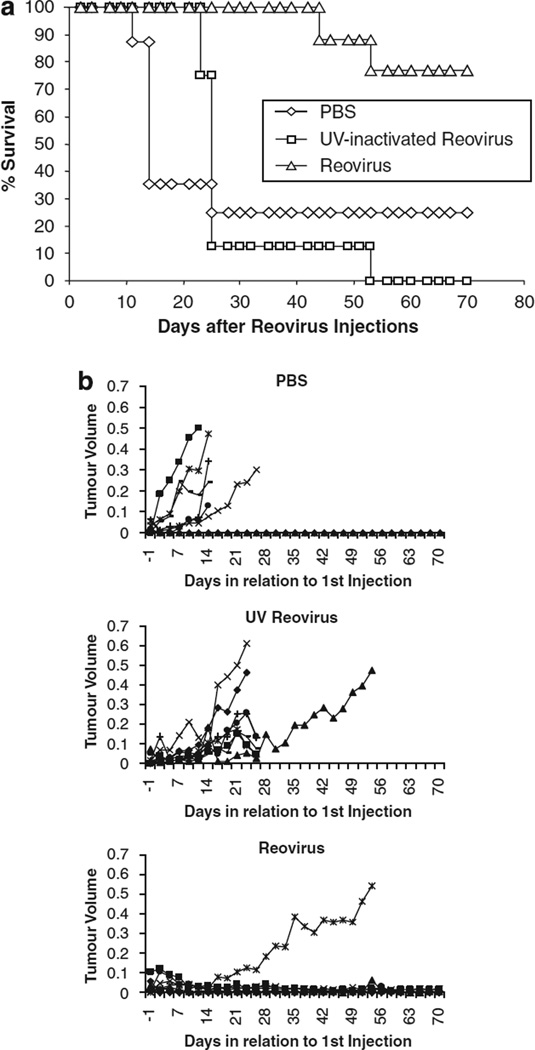
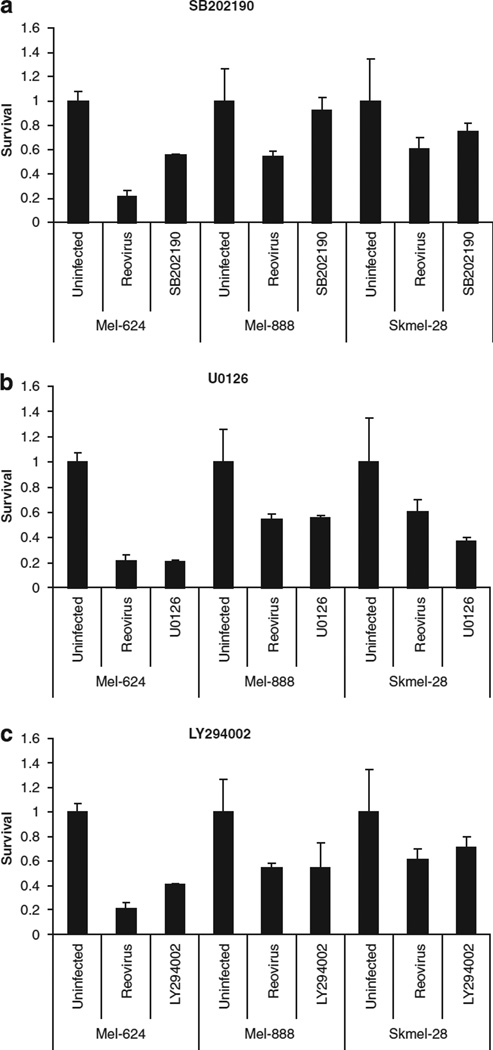
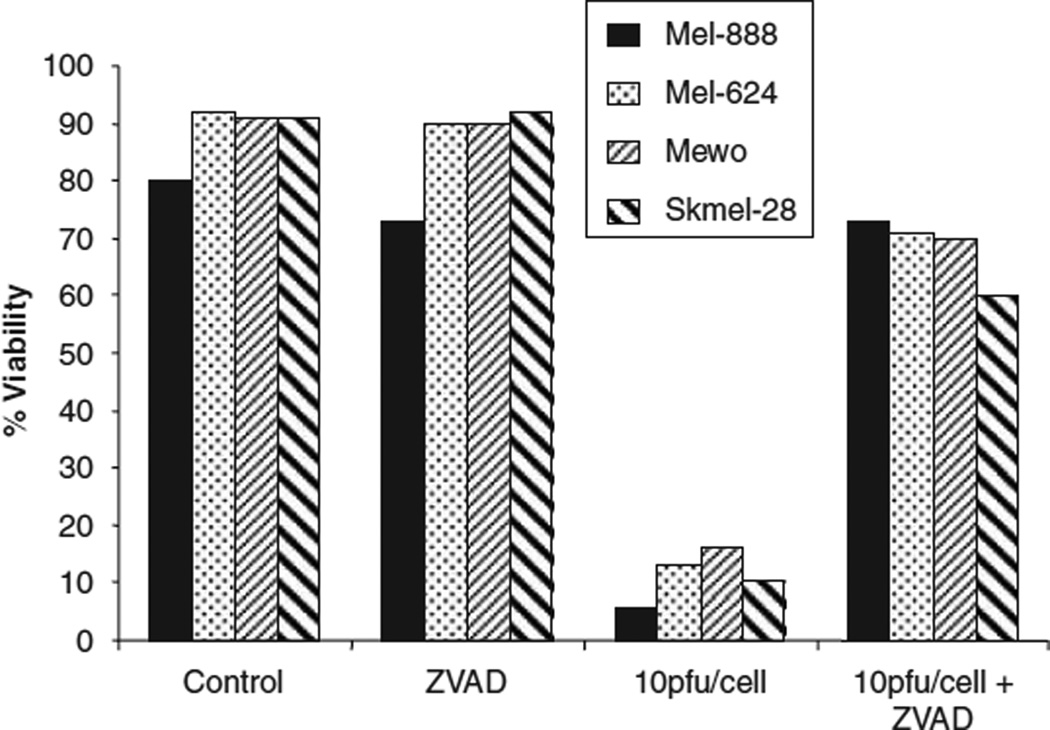
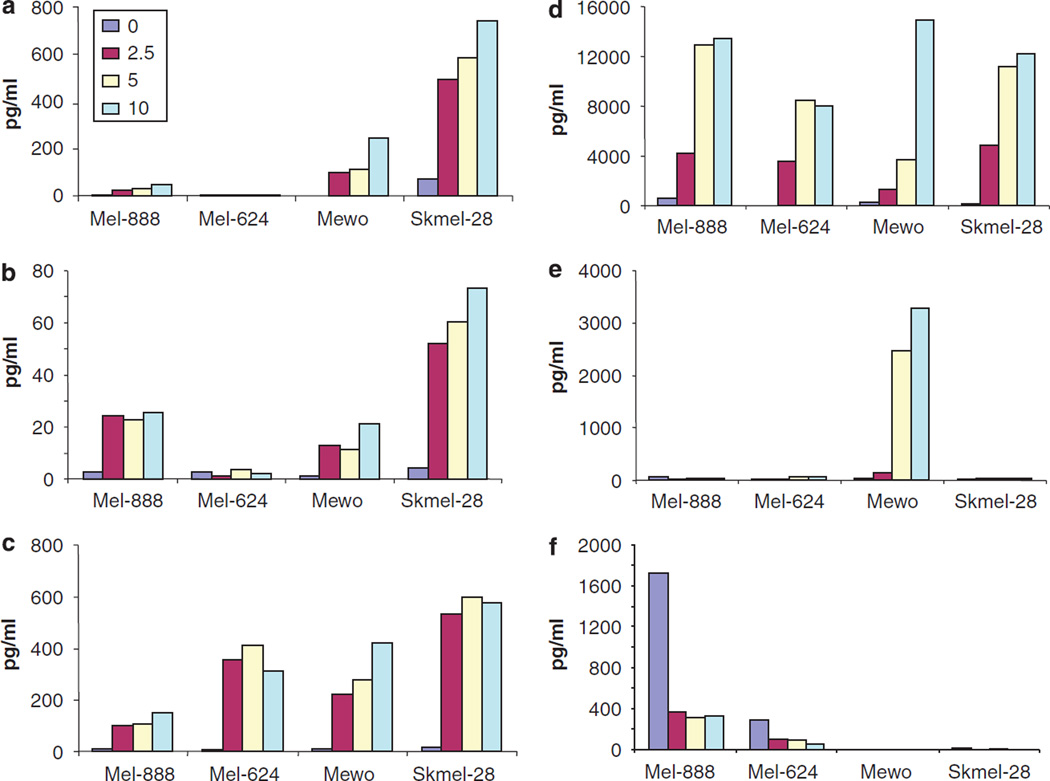
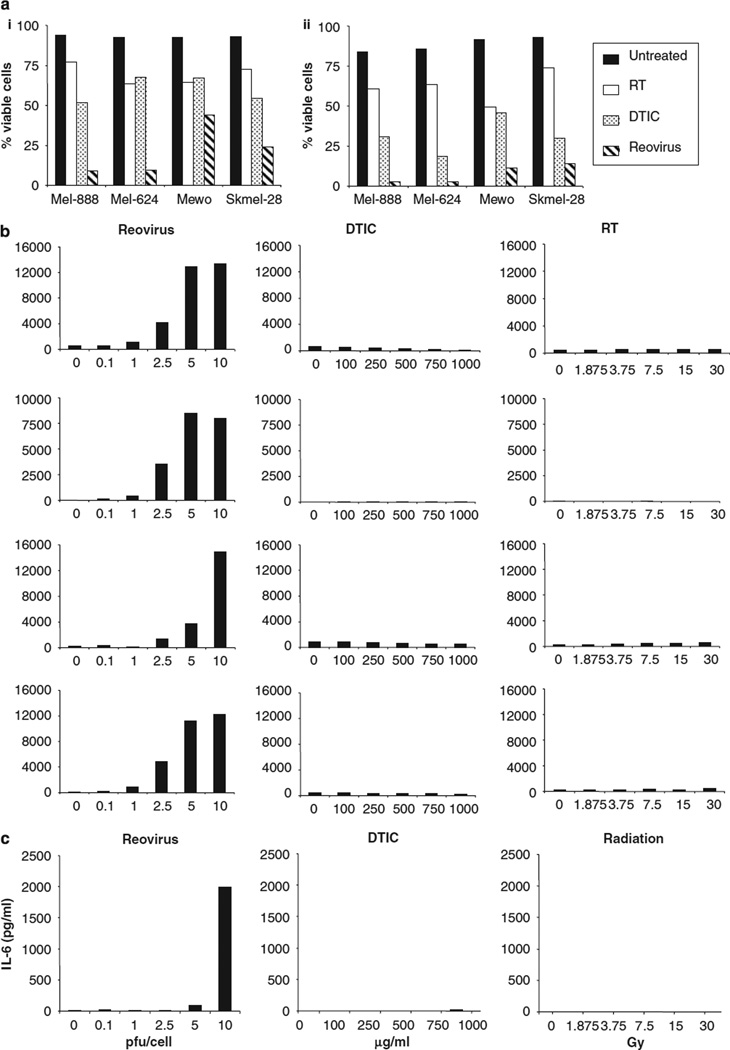
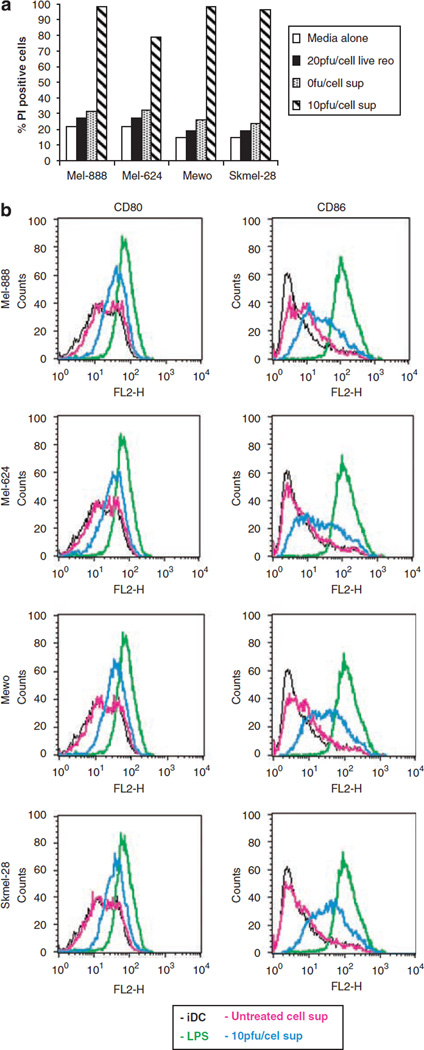
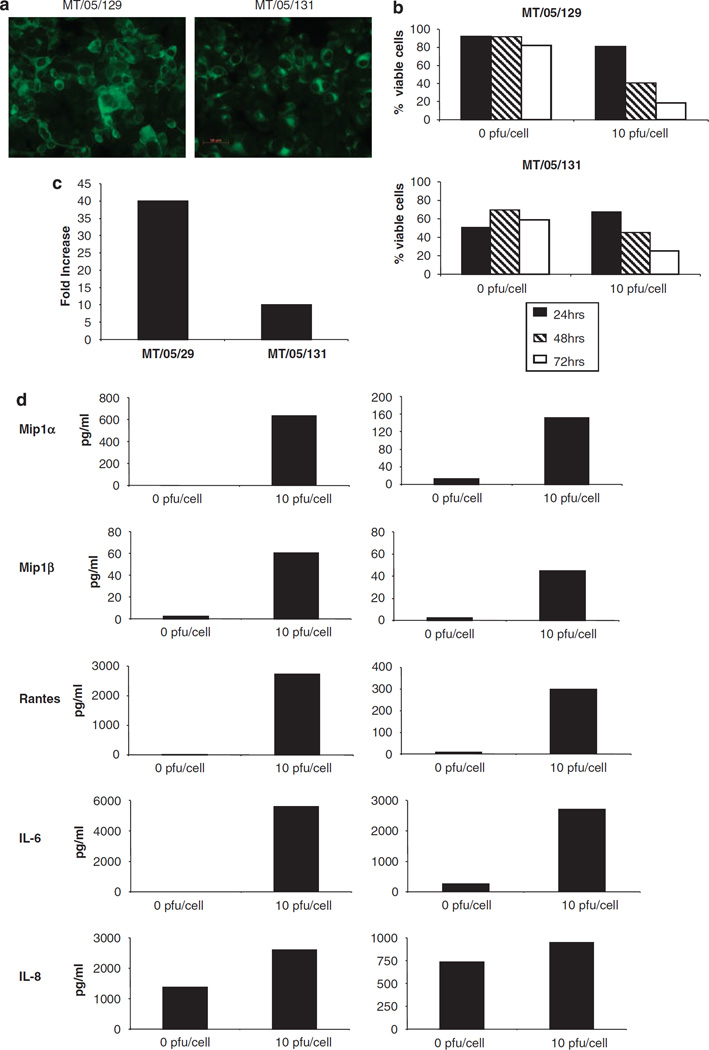
Reovirus is a promising unmodified double-stranded RNA (dsRNA) anti-cancer oncolytic virus, which is thought to specifically target cells with activated Ras. Although reovirus has been tested in a wide range of preclinical models and has entered early clinical trials, it has not previously been tested for the treatment of human melanoma. Here, we show that reovirus effectively kills and replicates in both human melanoma cell lines and freshly resected tumour; intratumoural injection also causes regression of melanoma in a xenograft in vivo model. Reovirus-induced melanoma death is blocked by caspase inhibition and is dependent on constituents of the Ras/RalGEF/p38 pathway. Reovirus melanoma killing is more potent than, and distinct from, chemotherapy or radiotherapy-induced cell death; a range of inflammatory cytokines and chemokines are released by infected tumour cells, while IL-10 secretion is abrogated. Furthermore, the inflammatory response generated by reovirus-infected tumour cells causes bystander toxicity against reovirus-resistant tumour cells and activates human myeloid dendritic cells (DC) in vitro. Hence, reovirus is suitable for clinical testing in melanoma, and may provide a useful danger signal to reverse the immunologically suppressive environment characteristic of this tumour.
Figures
References
-
- Selb B, Weber B. A study of human reovirus IgG and IgA antibodies by ELISA and western blot. J Virol Methods. 1994;47:15–25. - PubMed
-
- Coffey MC, Strong JE, Forsyth PA, Lee PW. Reovirus therapy of tumors with activated Ras pathway. Science. 1998;282:1332–1334. - PubMed
-
- Marcato P, Shmulevitz M, Lee PW. Connecting reovirus oncolysis and Ras signaling. Cell Cycle. 2005;4:556–559. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
