

CD39 and control of cellular immune responses
- PMID: 18404431
- PMCID: PMC2096766
- DOI: 10.1007/s11302-006-9050-y
CD39 and control of cellular immune responses
Abstract
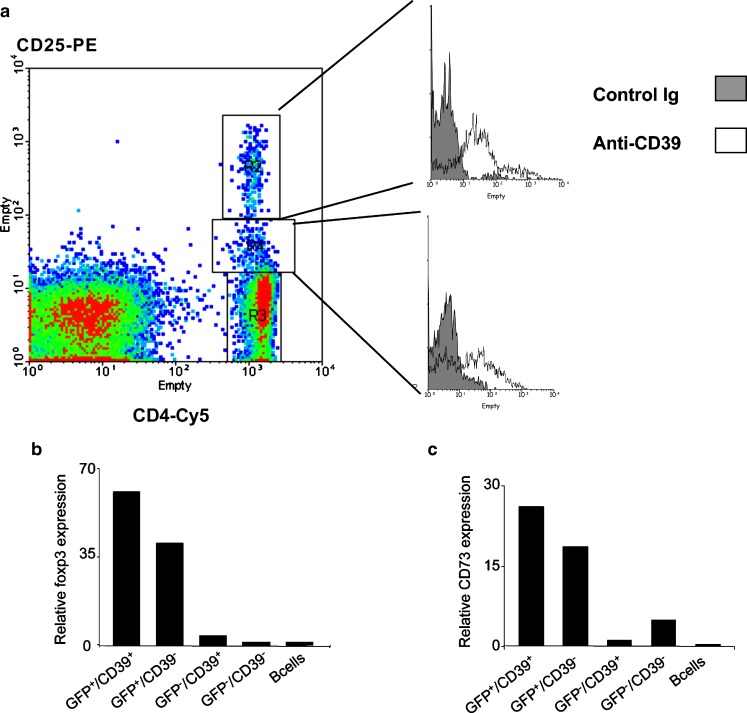
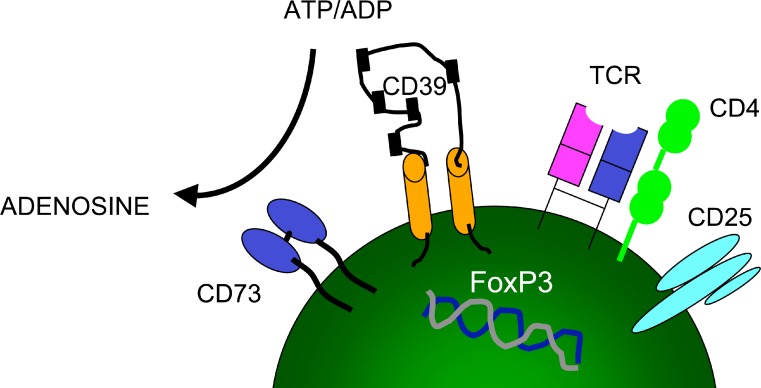
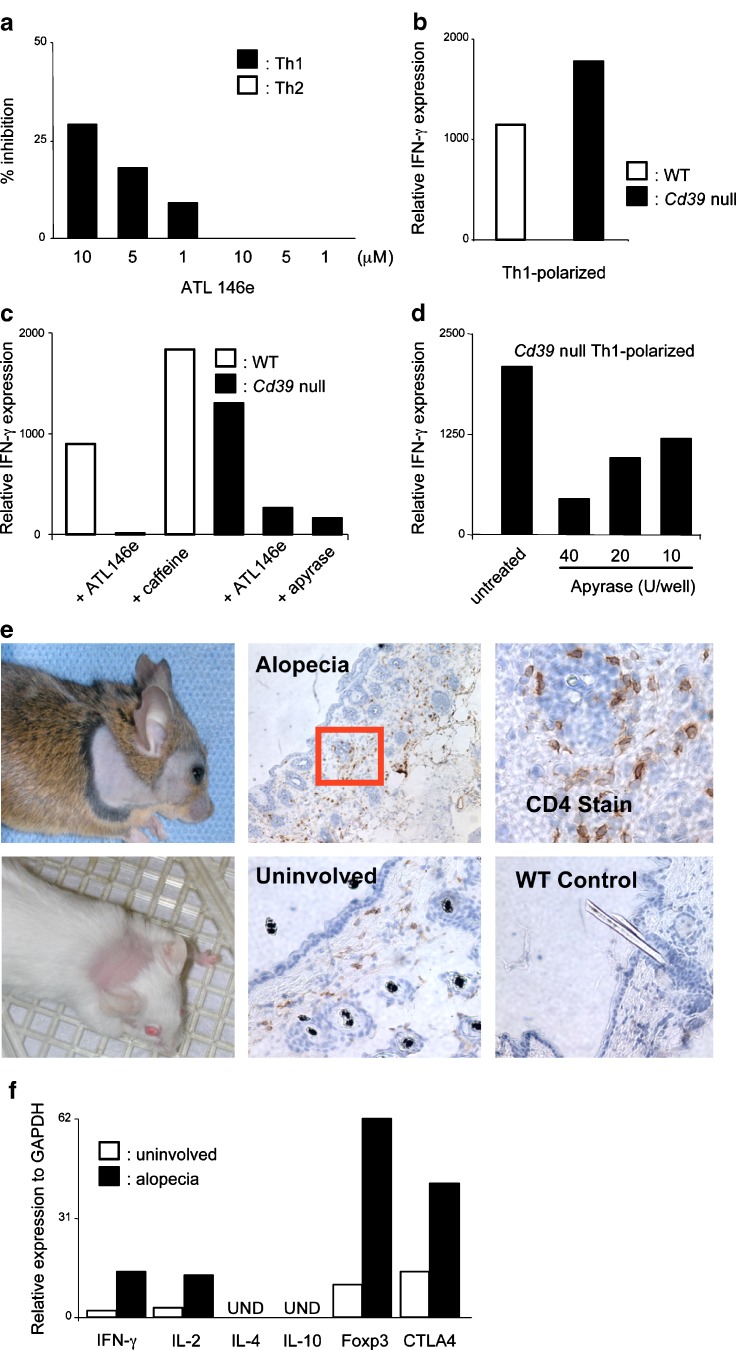
CD39 is the cell surface-located prototypic member of the ecto-nucleoside triphosphate diphosphohydrolase (E-NTPDase) family. Biological actions of CD39 are a consequence (at least in part) of the regulated phosphohydrolytic activity on extracellular nucleotides. This ecto-enzymatic cascade in tandem with CD73 (ecto-5'-nucleotidase) also generates adenosine and has major effects on both P2 and adenosine receptor signalling. Despite the early recognition of CD39 as a B lymphocyte activation marker, little is known of the role of CD39 in humoral or cellular immune responses. There is preliminary evidence to suggest that CD39 may impact upon antibody affinity maturation. Pericellular nucleotide/nucleoside fluxes caused by dendritic cell expressed CD39 are also involved in the recruitment, activation and polarization of naïve T cells. We have recently explored the patterns of CD39 expression and the functional role of this ecto-nucleotidase within quiescent and activated T cell subsets. Our data indicate that CD39, together with CD73, efficiently distinguishes T regulatory cells (Treg) from other resting or activated T cells in mice (and humans). Furthermore, CD39 serves as an integral component of the suppressive machinery of Treg, acting, at least in part, through the modulation of pericellular levels of adenosine. We have also shown that the coordinated regulation of CD39/CD73 expression and of the adenosine receptor A2A activates an immunoinhibitory loop that differentially regulates Th1 and Th2 responses. The in vivo relevance of this network is manifest in the phenotype of Cd39-null mice that spontaneously develop features of autoimmune diseases associated with Th1 immune deviation. These data indicate the potential of CD39 and modulated purinergic signalling in the co-ordination of immunoregulatory functions of dendritic and Treg cells. Our findings also suggest novel therapeutic strategies for immune-mediated diseases.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
