

The PI3K pathway regulates endochondral bone growth through control of hypertrophic chondrocyte differentiation
- PMID: 18405384
- PMCID: PMC2329617
- DOI: 10.1186/1471-213X-8-40
The PI3K pathway regulates endochondral bone growth through control of hypertrophic chondrocyte differentiation
Abstract
Background: The majority of our bones develop through the process of endochondral ossification that involves chondrocyte proliferation and hypertrophic differentiation in the cartilage growth plate. A large number of growth factors and hormones have been implicated in the regulation of growth plate biology, however, less is known about the intracellular signaling pathways involved. PI3K/Akt has been identified as a major regulator of cellular proliferation, differentiation and death in multiple cell types.
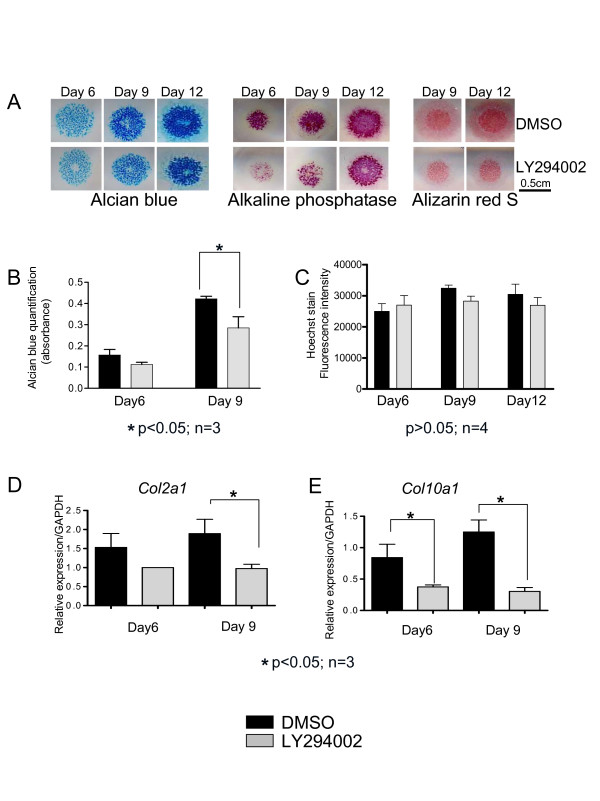
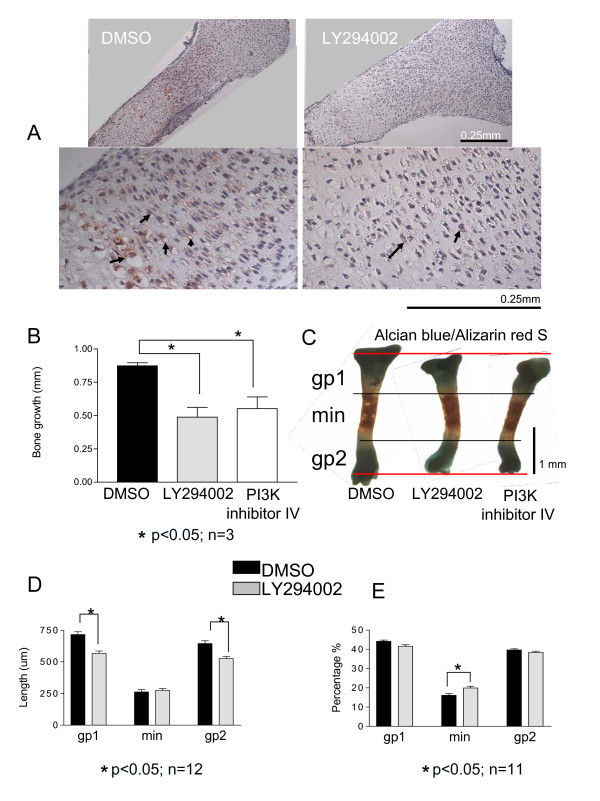
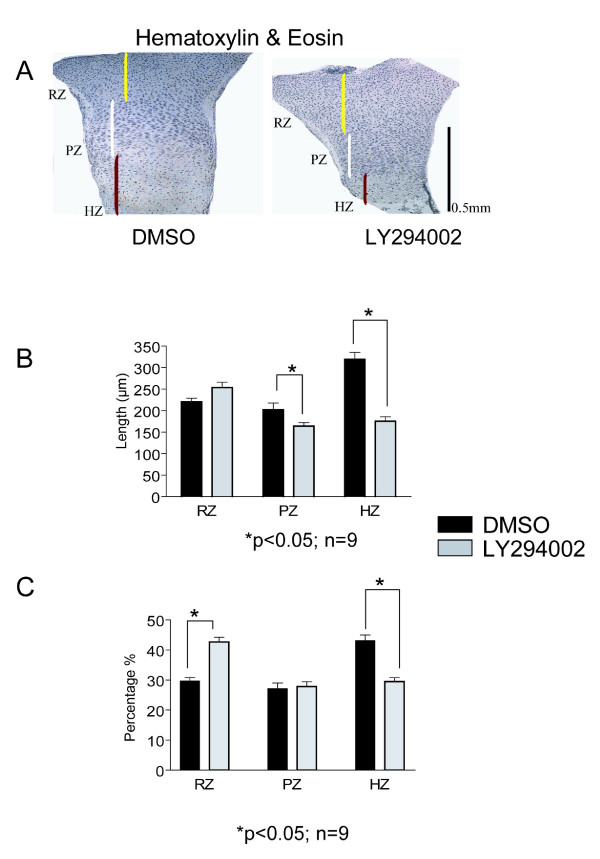
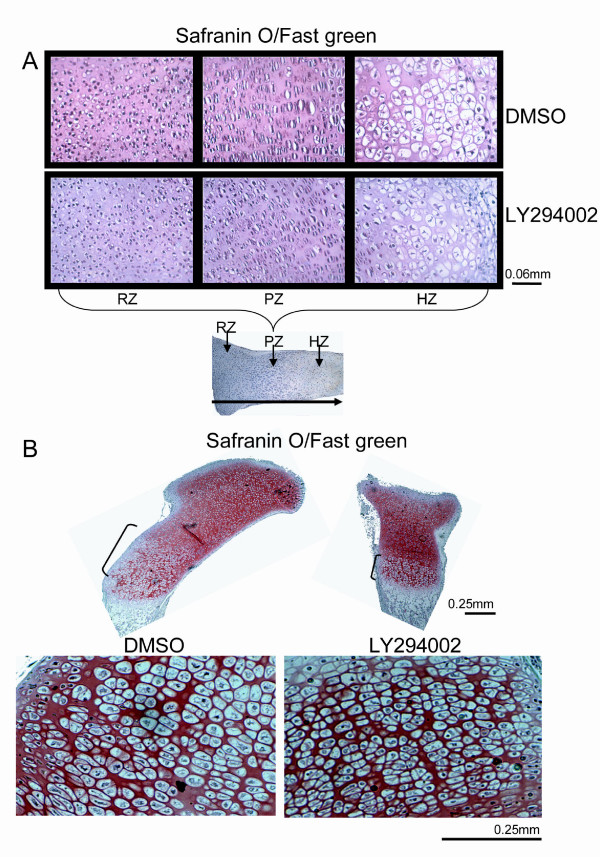
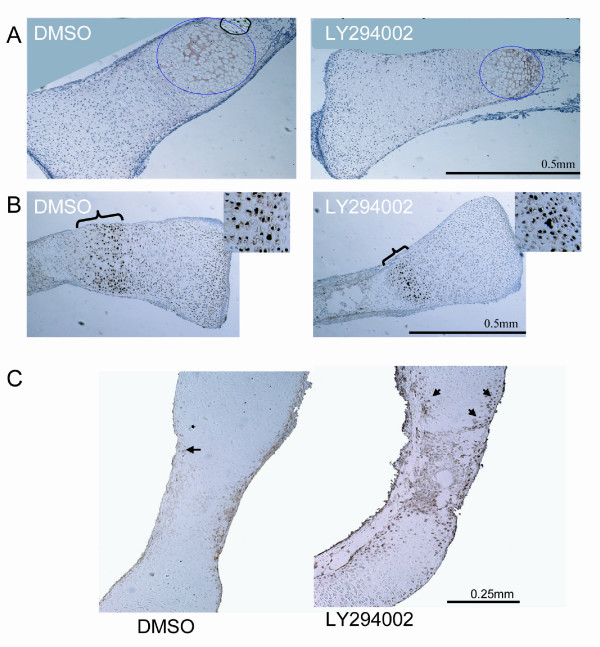
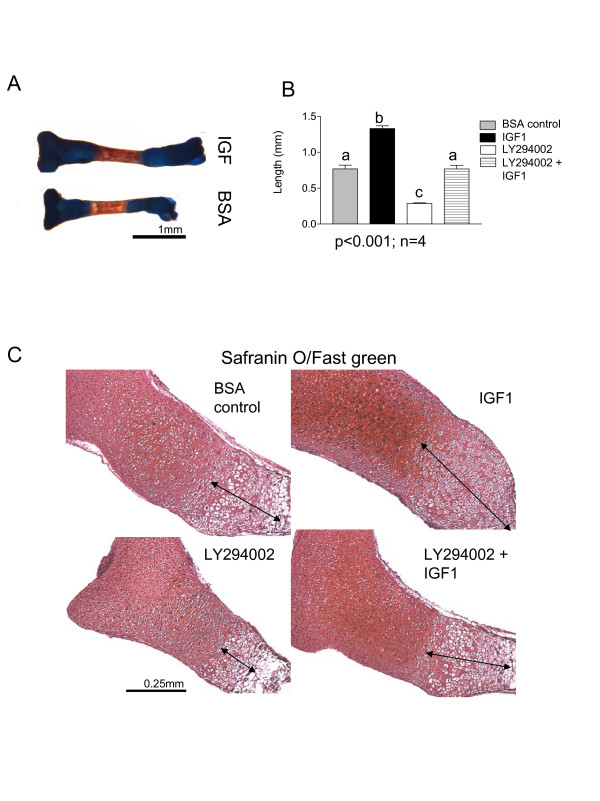
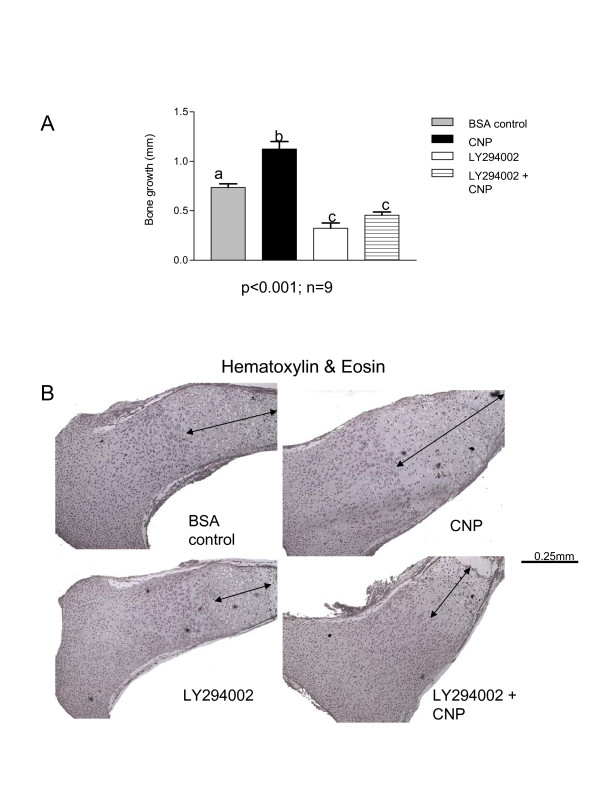
Results and discussion: Employing an organ culture system of embryonic mouse tibiae and LY294002, a pharmacological inhibitor of PI3K, we show that inhibition of the pathway results in significant growth reduction, demonstrating that PI3K is required for normal endochondral bone growth in vitro. PI3K inhibition reduces the length of the proliferating and particularly of the hypertrophic zone. Studies with organ cultures and primary chondrocytes in micromass culture show delayed hypertrophic differentiation of chondrocytes and increased apoptosis in the presence of LY294002. Surprisingly, PI3K inhibition had no strong effect on IGF1-induced bone growth, but partially blocked the anabolic effects of C-type natriuretic peptide.
Conclusion: Our data demonstrate an essential role of PI3K signaling in chondrocyte differentiation and as a consequence of this, in the endochondral bone growth process.
Figures
Similar articles
-
Regulation of gene expression by PI3K in mouse growth plate chondrocytes.PLoS One. 2010 Jan 25;5(1):e8866. doi: 10.1371/journal.pone.0008866. PLoS One. 2010. PMID: 20111593 Free PMC article.
-
8-Nitro-cGMP promotes bone growth through expansion of growth plate cartilage.Free Radic Biol Med. 2017 Sep;110:63-71. doi: 10.1016/j.freeradbiomed.2017.05.022. Epub 2017 May 27. Free Radic Biol Med. 2017. PMID: 28559051
-
Liver X Receptor activation delays chondrocyte hypertrophy during endochondral bone growth.Osteoarthritis Cartilage. 2014 Jul;22(7):996-1006. doi: 10.1016/j.joca.2014.05.003. Epub 2014 May 20. Osteoarthritis Cartilage. 2014. PMID: 24852699
-
Chondrocyte hypertrophy in skeletal development, growth, and disease.Birth Defects Res C Embryo Today. 2014 Mar;102(1):74-82. doi: 10.1002/bdrc.21062. Birth Defects Res C Embryo Today. 2014. PMID: 24677724 Review.
-
Expression of bone-specific genes by hypertrophic chondrocytes: implication of the complex functions of the hypertrophic chondrocyte during endochondral bone development.J Cell Biochem. 1996 Jul;62(1):1-9. doi: 10.1002/(SICI)1097-4644(199607)62:1%3C1::AID-JCB1%3E3.0.CO;2-X. J Cell Biochem. 1996. PMID: 8836870 Review.
Cited by
-
Vinculin functions as regulator of chondrogenesis.J Biol Chem. 2012 May 4;287(19):15760-75. doi: 10.1074/jbc.M111.308072. Epub 2012 Mar 13. J Biol Chem. 2012. PMID: 22416133 Free PMC article.
-
Microarray analysis identification of key pathways and interaction network of differential gene expressions during osteogenic differentiation.Hum Genomics. 2020 Nov 25;14(1):43. doi: 10.1186/s40246-020-00293-1. Hum Genomics. 2020. PMID: 33234152 Free PMC article.
-
Long-term dynamic compression enhancement TGF-β3-induced chondrogenesis in bovine stem cells: a gene expression analysis.BMC Genom Data. 2021 Mar 20;22(1):13. doi: 10.1186/s12863-021-00967-2. BMC Genom Data. 2021. PMID: 33743603 Free PMC article.
-
Skeletal defects and bone metabolism in Noonan, Costello and cardio-facio-cutaneous syndromes.Front Endocrinol (Lausanne). 2023 Oct 27;14:1231828. doi: 10.3389/fendo.2023.1231828. eCollection 2023. Front Endocrinol (Lausanne). 2023. PMID: 37964950 Free PMC article. Review.
-
Low-dose IL-34 has no effect on osteoclastogenesis but promotes osteogenesis of hBMSCs partly via activation of the PI3K/AKT and ERK signaling pathways.Stem Cell Res Ther. 2021 May 4;12(1):268. doi: 10.1186/s13287-021-02263-3. Stem Cell Res Ther. 2021. PMID: 33947456 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
