

Tonic and stimulus-evoked nitric oxide production in the mouse olfactory bulb
- PMID: 18407420
- PMCID: PMC2475540
- DOI: 10.1016/j.neuroscience.2008.03.003
Tonic and stimulus-evoked nitric oxide production in the mouse olfactory bulb
Abstract
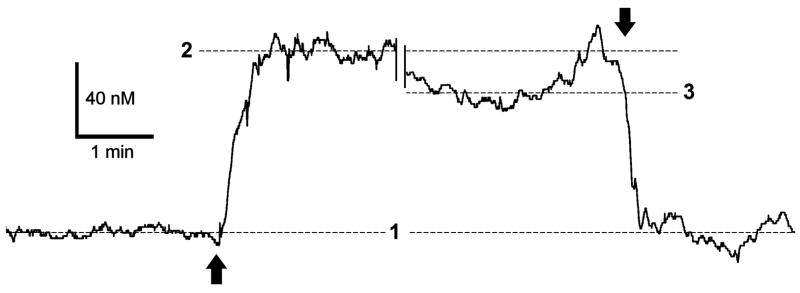
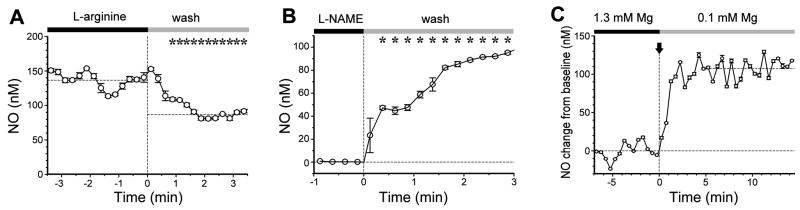
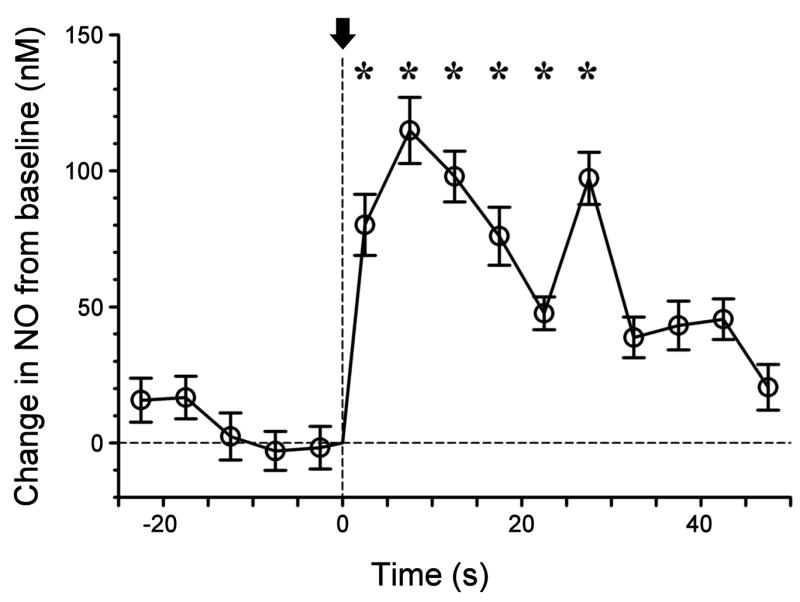
Nitric oxide (NO) has been long assumed to play a key role in mammalian olfaction. This was based largely on circumstantial evidence, i.e. prominent staining for nitric oxide synthase (NOS) and cyclic guanosine 3',5'-cyclic monophosphate (cGMP) or soluble guanylyl cyclase, an effector enzyme activated by NO, in local interneurons of the olfactory bulb. Here we employ innovative custom-fabricated NO micro-sensors to obtain the first direct, time-resolved measurements of NO signaling in the olfactory bulb. In 400 microm thick mouse olfactory bulb slices, we detected a steady average basal level of 87 nM NO in the extracellular space of mitral or granule cell layers. This NO 'tone' was sensitive to NOS substrate manipulation (200 microM L-arginine, 2 mM N(G)-nitro-L-arginine methyl ester) and Mg(2+) modulation of N-methyl-D-aspartate (NMDA) receptor conductance. Electrical stimulation of olfactory nerve fibers evoked transient (peak at 10 s) increments in NO levels 90-100 nM above baseline. In the anesthetized mouse, NO micro-sensors inserted into the granule cell layer detected NO transients averaging 55 nM in amplitude and peaking at 3.4 s after onset of a 5 s odorant stimulation. These findings suggest dual roles for NO signaling in the olfactory bulb: tonic inhibitory control of principal neurons, and regulation of circuit dynamics during odor information processing.
Figures

References
-
- Ahern GP, Klyachko VA, Jackson MB. cGMP and S-nitrosylation: two routes for modulation of neuronal excitability by NO. Trends Neurosci. 2002;25:510–517. - PubMed
-
- Allen BW, Piantadosi CA, Coury LAJ. Electrode materials for nitric oxide detection. Nitric Oxide: Biology and Chemistry. 2000;4:75–84. - PubMed
-
- Alonso JR, Porteros A, Crespo C, Arevalo R, Brinon JG, Weruaga E, Aijon J. Chemical anatomy of the macaque monkey olfactory bulb: NADPH-diaphorase/nitric oxide synthase activity. J Comp Neurol. 1998;402:419–434. - PubMed
-
- Bohlen HG. Mechanism of increased vessel wall nitric oxide concentrations during intestinal absorption. Am J Physiol Heart Circ Physiol. 1998;275:H542–H550. - PubMed
-
- Bredt DS, Glatt CE, Hwang PM, Fotuhi M, Dawson TM, Snyder SH. Nitric oxide synthase protein and mRNA are discretely localized in neuronal populations of the mammalian CNS together with NADPH diaphorase. Neuron. 1991;7:615–624. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
