

The peptidoglycan of stationary-phase Mycobacterium tuberculosis predominantly contains cross-links generated by L,D-transpeptidation
- PMID: 18408028
- PMCID: PMC2446752
- DOI: 10.1128/JB.00239-08
The peptidoglycan of stationary-phase Mycobacterium tuberculosis predominantly contains cross-links generated by L,D-transpeptidation
Abstract
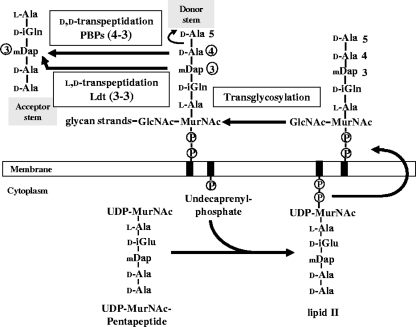
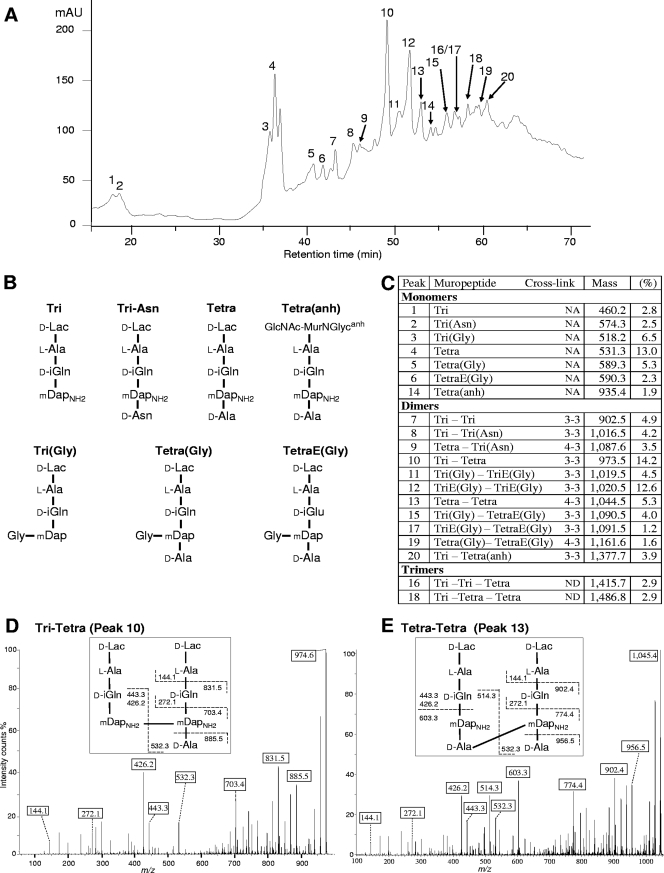
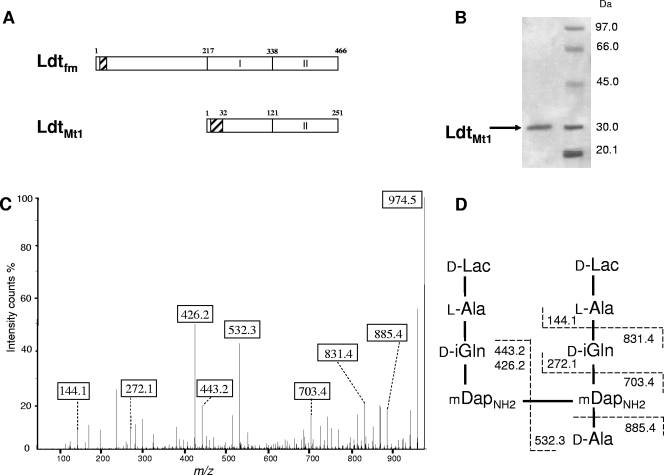
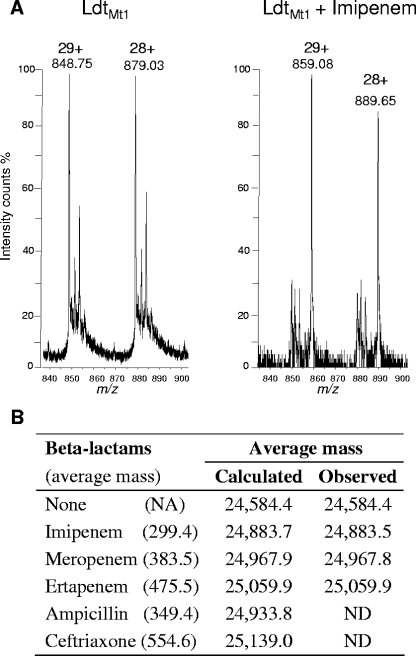
Our understanding of the mechanisms used by Mycobacterium tuberculosis to persist in a "dormant" state is essential to the development of therapies effective in sterilizing tissues. Gene expression profiling in model systems has revealed a complex adaptive response thought to endow M. tuberculosis with the capacity to survive several months of combinatorial antibiotic treatment. We show here that this adaptive response may involve remodeling of the peptidoglycan network by substitution of 4-->3 cross-links generated by the D,D-transpeptidase activity of penicillin-binding proteins by 3-->3 cross-links generated by a transpeptidase of L,D specificity. A candidate gene, previously shown to be upregulated upon nutrient starvation, was found to encode an L,D-transpeptidase active in the formation of 3-->3 cross-links. The enzyme, Ldt(Mt1), was inactivated by carbapenems, a class of beta-lactam antibiotics that are poorly hydrolyzed by the M. tuberculosis beta-lactamases. Ldt(Mt1) and carbapenems may therefore represent a target and a drug family relevant to the eradication of persistent M. tuberculosis.
Figures
References
-
- Amrein, K. E., B. Takacs, M. Stieger, J. Molnos, N. A. Flint, and P. Burn. 1995. Purification and characterization of recombinant human p50csk protein-tyrosine kinase from an Escherichia coli expression system overproducing the bacterial chaperones GroES and GroEL. Proc. Natl. Acad. Sci. USA 921048-1052. - PMC - PubMed
-
- Arbeloa, A., J. E. Hugonnet, A. C. Sentilhes, N. Josseaume, L. Dubost, C. Monsempes, D. Blanot, J. P. Brouard, and M. Arthur. 2004. Synthesis of mosaic peptidoglycan cross-bridges by hybrid peptidoglycan assembly pathways in gram-positive bacteria. J. Biol. Chem. 27941546-41556. - PubMed
-
- Auger, G., J. van Heijenoort, D. Mengin-Lecreulx, and D. Blanot. 2003. A MurG assay which utilizes a synthetic analogue of lipid I. FEMS Microbiol. Lett. 219115-119. - PubMed
-
- Betts, J. C., P. T. Lukey, L. C. Robb, R. A. McAdam, and K. Duncan. 2002. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 43717-731. - PubMed
-
- Biarrotte-Sorin, S., J. E. Hugonnet, V. Delfosse, J. L. Mainardi, L. Gutmann, M. Arthur, and C. Mayer. 2006. Crystal structure of a novel beta-lactam-insensitive peptidoglycan transpeptidase. J. Mol. Biol. 359533-538. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
