

HD-ZIP III activity is modulated by competitive inhibitors via a feedback loop in Arabidopsis shoot apical meristem development
- PMID: 18408069
- PMCID: PMC2390745
- DOI: 10.1105/tpc.107.057448
HD-ZIP III activity is modulated by competitive inhibitors via a feedback loop in Arabidopsis shoot apical meristem development
Abstract
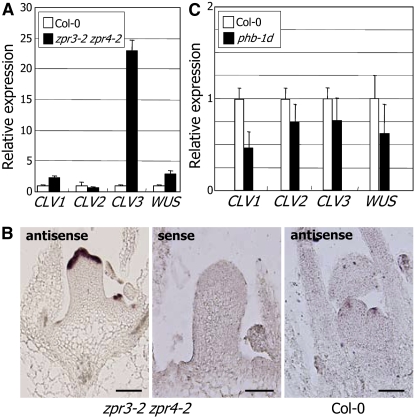
Shoot apical meristem (SAM) development is coordinately regulated by two interdependent signaling events: one maintaining stem cell identity and the other governing the initiation of lateral organs from the flanks of the SAM. The signaling networks involved in this process are interconnected and are regulated by multiple molecular mechanisms. Class III homeodomain-leucine zipper (HD-ZIP III) proteins are the most extensively studied transcription factors involved in this regulation. However, how different signals are integrated to maintain stem cell identity and to pattern lateral organ polarity remains unclear. Here, we demonstrated that a small ZIP protein, ZPR3, and its functionally redundant homolog, ZPR4, negatively regulate the HD-ZIP III activity in SAM development. ZPR3 directly interacts with PHABULOSA (PHB) and other HD-ZIP III proteins via the ZIP motifs and forms nonfunctional heterodimers. Accordingly, a double mutant, zpr3-2 zpr4-2, exhibits an altered SAM activity with abnormal stem cell maintenance. However, the mutant displays normal patterning of leaf polarity. In addition, we show that PHB positively regulates ZPR3 expression. We therefore propose that HD-ZIP III activity in regulating SAM development is modulated by, among other things, a feedback loop involving the competitive inhibitors ZPR3 and ZPR4.
Figures







References
-
- Baima, S., Nobili, F., Sessa, G., Lucchetti, S., Ruberti, I., and Morelli, G. (1995). The expression of the ATHB-8 homeobox gene is restricted to provascular cells in Arabidopsis thaliana. Development 121 4171–4182. - PubMed
-
- Benezra, R., Davis, R.L., Lockshon, D., Turner, D.L., and Weintraub, H. (1990). The protein ID: A negative regulator of helix-loop-helix DNA binding proteins. Cell 61 49–59. - PubMed
-
- Bowman, J.L. (2004). Class III HD-ZIP gene regulation, the golden fleece of ARGONAUTE activity? Bioessays 26 938–942. - PubMed
-
- Brand, U., Fletcher, J.C., Hobe, M., Meyerowitz, E.M., and Simon, R. (2000). Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289 617–619. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
