

The HSA domain binds nuclear actin-related proteins to regulate chromatin-remodeling ATPases
- PMID: 18408732
- PMCID: PMC2810487
- DOI: 10.1038/nsmb.1403
The HSA domain binds nuclear actin-related proteins to regulate chromatin-remodeling ATPases
Abstract
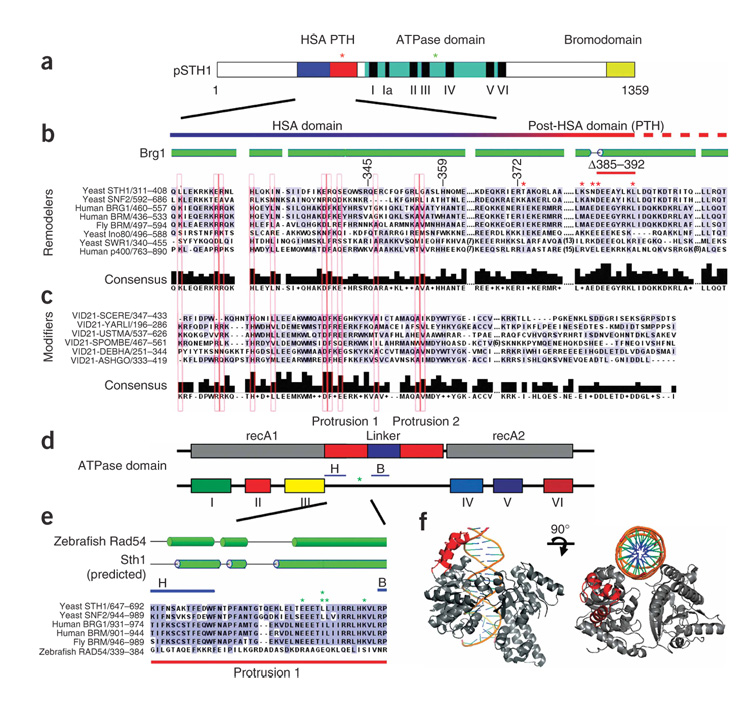
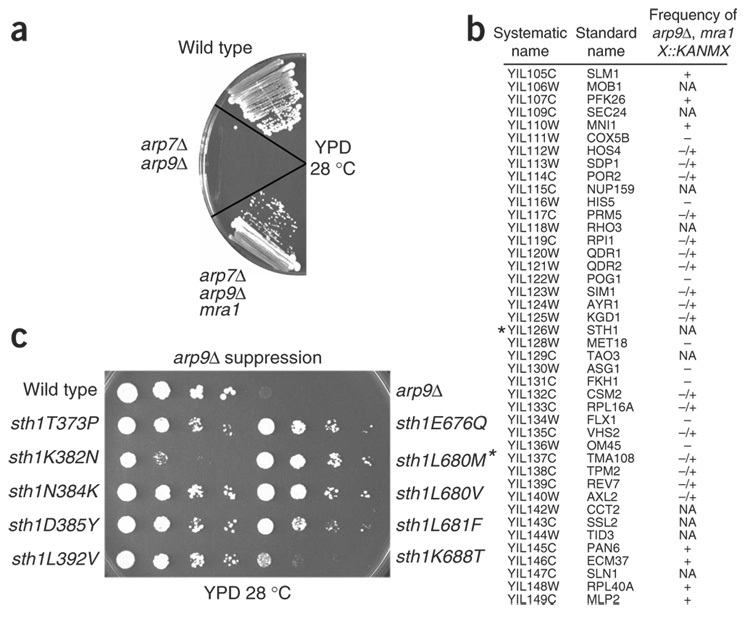
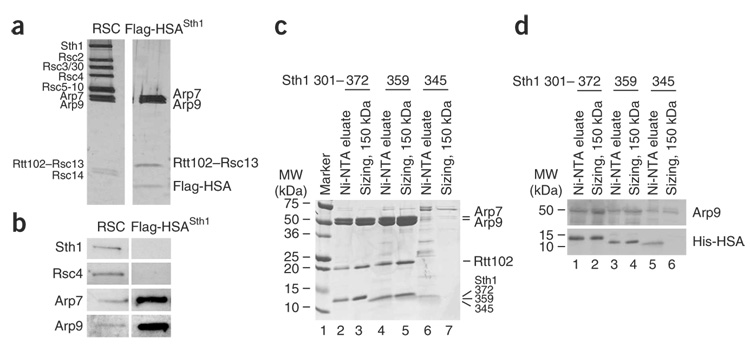
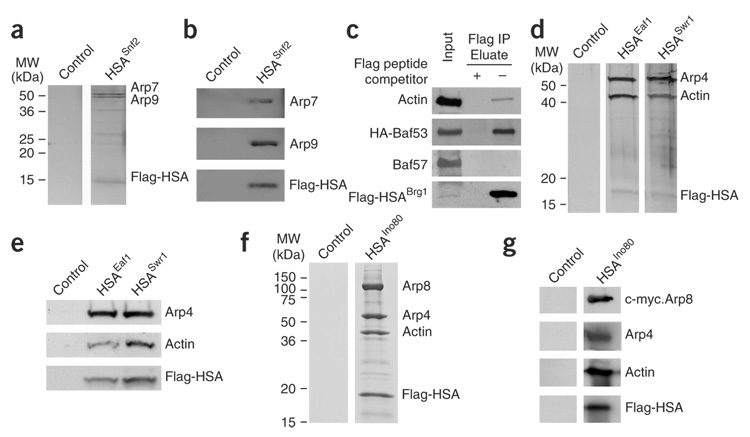
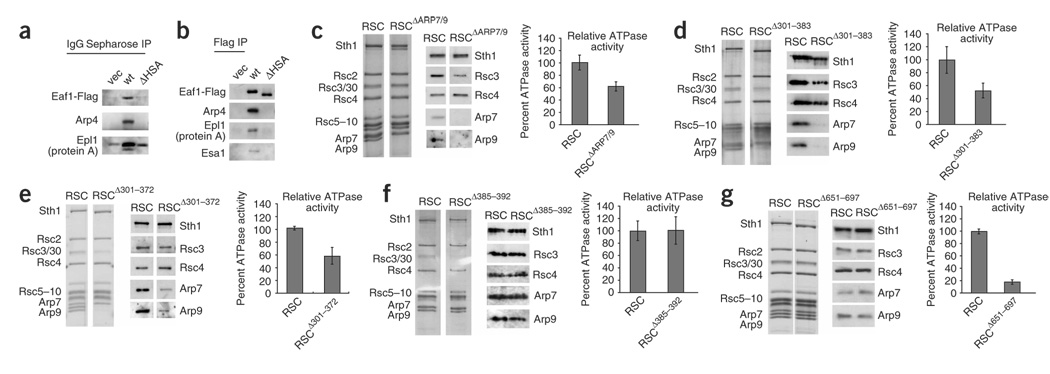
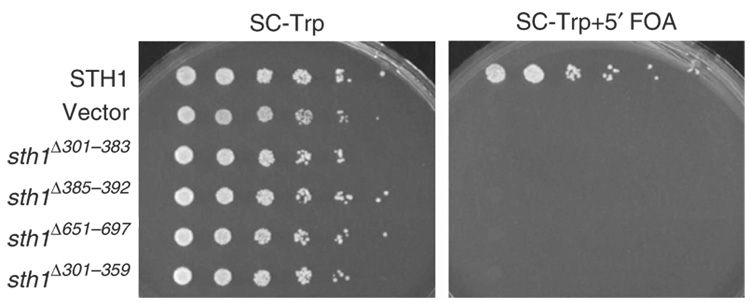
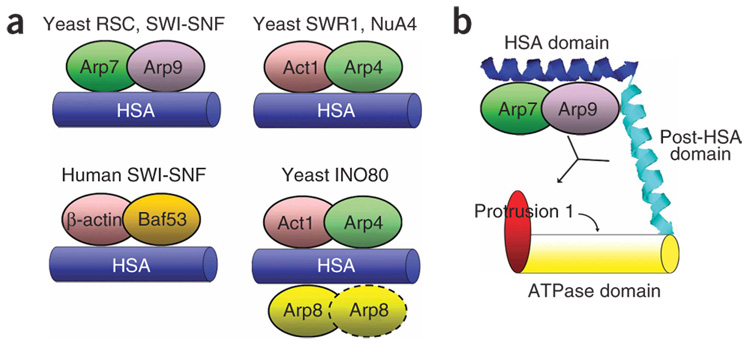
We identify the helicase-SANT-associated (HSA) domain as the primary binding platform for nuclear actin-related proteins (ARPs) and actin. Individual HSA domains from chromatin remodelers (RSC, yeast SWI-SNF, human SWI-SNF, SWR1 and INO80) or modifiers (NuA4) reconstitute their respective ARP-ARP or ARP-actin modules. In RSC, the HSA domain resides on the catalytic ATPase subunit Sth1. The Sth1 HSA is essential in vivo, and its omission causes the specific loss of ARPs and a moderate reduction in ATPase activity. Genetic selections for arp suppressors yielded specific gain-of-function mutations in two new domains in Sth1, the post-HSA domain and protrusion 1, which are essential for RSC function in vivo but not ARP association. Taken together, we define the role of the HSA domain and provide evidence for a regulatory relationship involving the ARP-HSA module and two new functional domains conserved in remodeler ATPases that contain ARPs.
Figures
Comment in
-
New clues to actin function in chromatin regulation.Nat Struct Mol Biol. 2008 May;15(5):432-3. doi: 10.1038/nsmb0508-432. Nat Struct Mol Biol. 2008. PMID: 18461045 No abstract available.
Similar articles
-
Structural insights into assembly and function of the RSC chromatin remodeling complex.Nat Struct Mol Biol. 2021 Jan;28(1):71-80. doi: 10.1038/s41594-020-00528-8. Epub 2020 Dec 7. Nat Struct Mol Biol. 2021. PMID: 33288924 Free PMC article.
-
Structure of an actin-related subcomplex of the SWI/SNF chromatin remodeler.Proc Natl Acad Sci U S A. 2013 Feb 26;110(9):3345-50. doi: 10.1073/pnas.1215379110. Epub 2013 Feb 11. Proc Natl Acad Sci U S A. 2013. PMID: 23401505 Free PMC article.
-
Actin-related proteins regulate the RSC chromatin remodeler by weakening intramolecular interactions of the Sth1 ATPase.Commun Biol. 2018;1:1. doi: 10.1038/s42003-017-0002-6. Epub 2018 Jan 22. Commun Biol. 2018. PMID: 29809203 Free PMC article.
-
INO80 subfamily of chromatin remodeling complexes.Mutat Res. 2007 May 1;618(1-2):18-29. doi: 10.1016/j.mrfmmm.2006.10.006. Epub 2007 Jan 21. Mutat Res. 2007. PMID: 17316710 Free PMC article. Review.
-
Actin-related proteins (Arps): conformational switches for chromatin-remodeling machines?Bioessays. 2000 Jul;22(7):666-72. doi: 10.1002/1521-1878(200007)22:7<666::AID-BIES9>3.0.CO;2-Y. Bioessays. 2000. PMID: 10878579 Review.
Cited by
-
The INO80 Complex Requires the Arp5-Ies6 Subcomplex for Chromatin Remodeling and Metabolic Regulation.Mol Cell Biol. 2016 Jan 11;36(6):979-91. doi: 10.1128/MCB.00801-15. Mol Cell Biol. 2016. PMID: 26755556 Free PMC article.
-
ACTL6A promotes repair of cisplatin-induced DNA damage, a new mechanism of platinum resistance in cancer.Proc Natl Acad Sci U S A. 2021 Jan 19;118(3):e2015808118. doi: 10.1073/pnas.2015808118. Proc Natl Acad Sci U S A. 2021. PMID: 33408251 Free PMC article.
-
Arabidopsis actin-related protein ARP5 in multicellular development and DNA repair.Dev Biol. 2009 Nov 1;335(1):22-32. doi: 10.1016/j.ydbio.2009.08.006. Epub 2009 Aug 11. Dev Biol. 2009. PMID: 19679120 Free PMC article.
-
Plant BCL-DOMAIN HOMOLOG proteins play a conserved role in SWI/SNF complex stability.Proc Natl Acad Sci U S A. 2025 Jan 21;122(3):e2413346122. doi: 10.1073/pnas.2413346122. Epub 2025 Jan 17. Proc Natl Acad Sci U S A. 2025. PMID: 39823297 Free PMC article.
-
Energy-driven genome regulation by ATP-dependent chromatin remodellers.Nat Rev Mol Cell Biol. 2024 Apr;25(4):309-332. doi: 10.1038/s41580-023-00683-y. Epub 2023 Dec 11. Nat Rev Mol Cell Biol. 2024. PMID: 38081975 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
