

Structure of Ca2+-bound S100A4 and its interaction with peptides derived from nonmuscle myosin-IIA
- PMID: 18410126
- PMCID: PMC2633413
- DOI: 10.1021/bi702537s
Structure of Ca2+-bound S100A4 and its interaction with peptides derived from nonmuscle myosin-IIA
Abstract
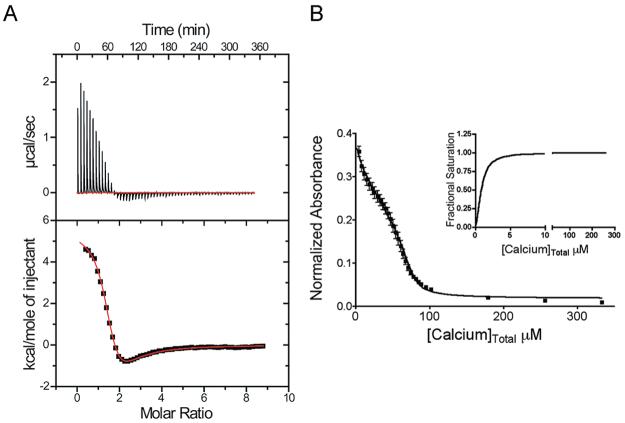
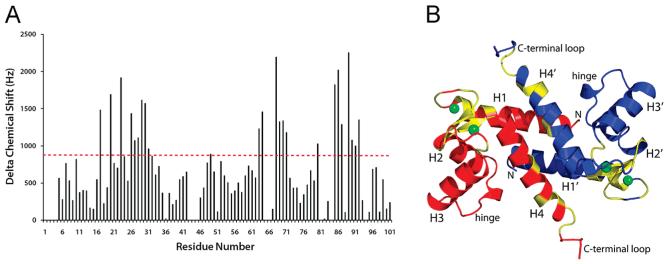
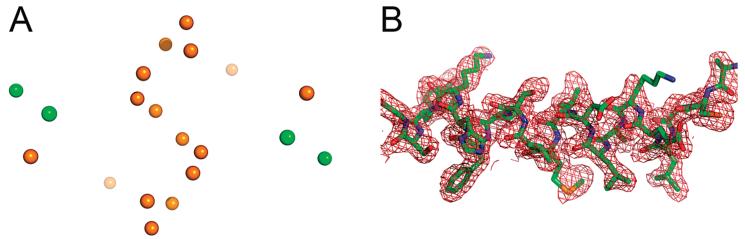
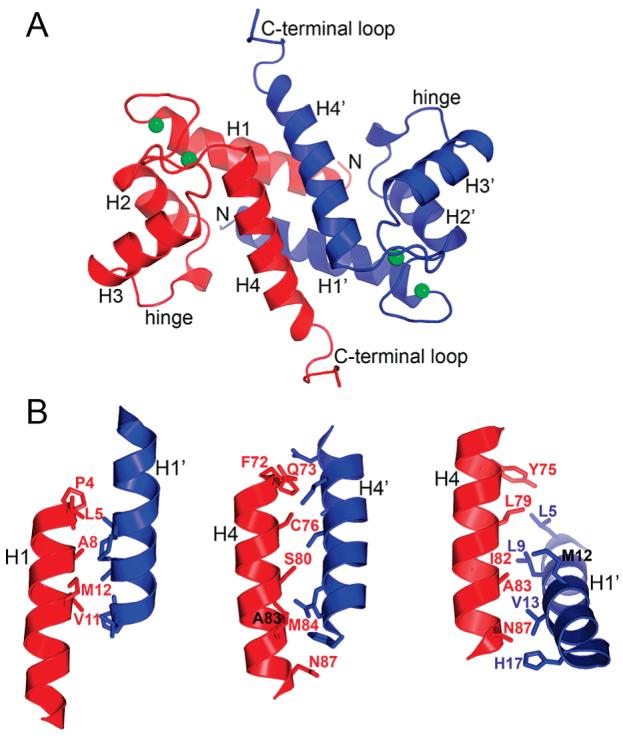
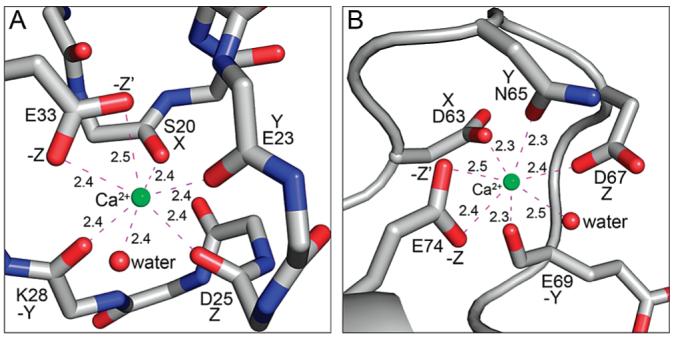
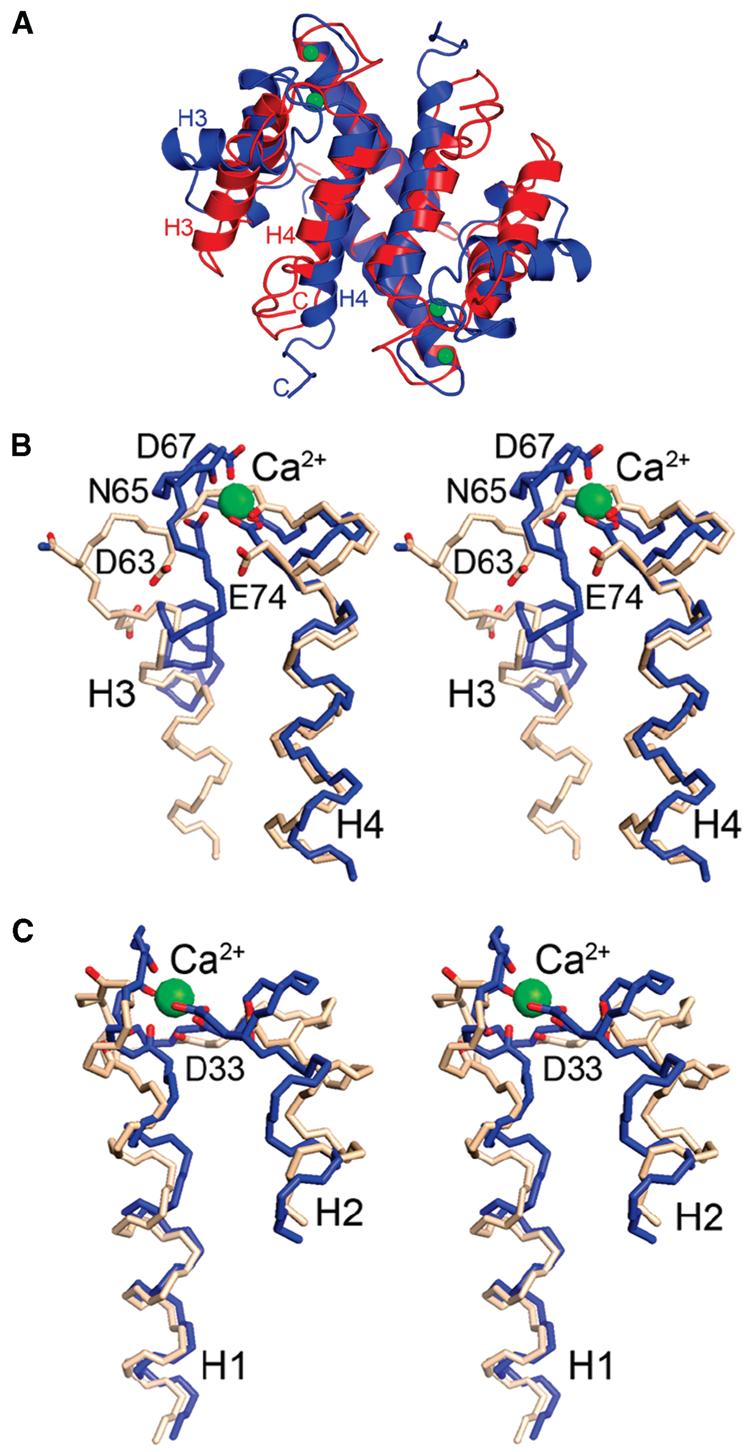
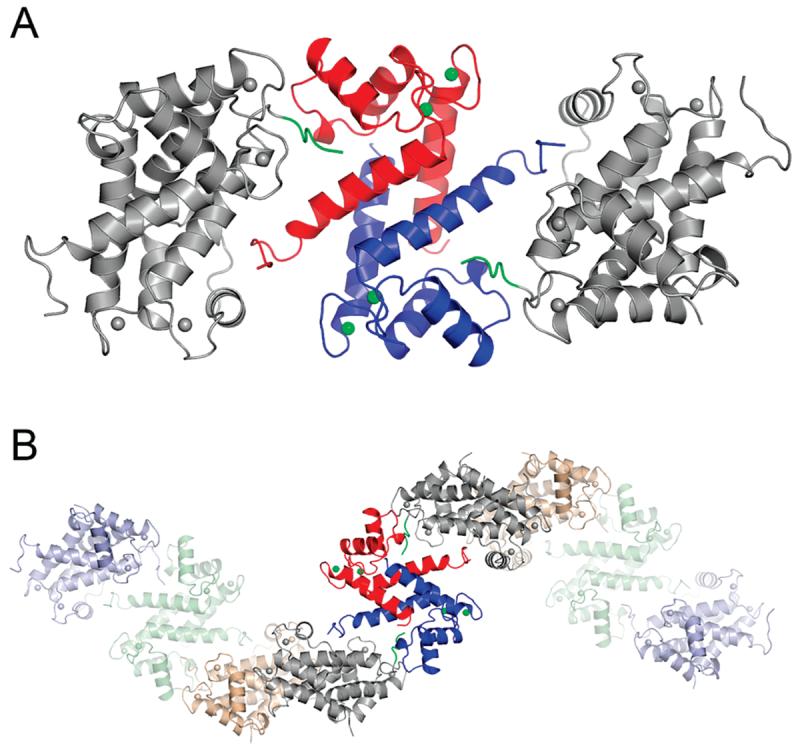
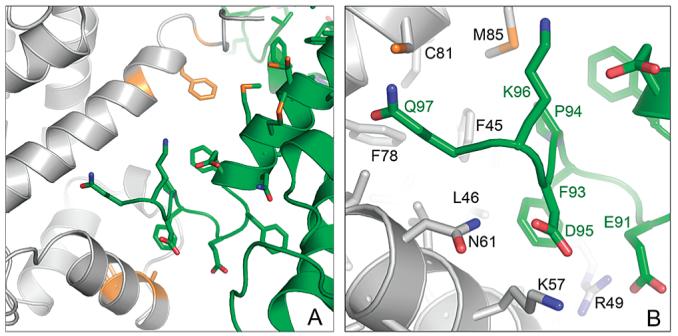
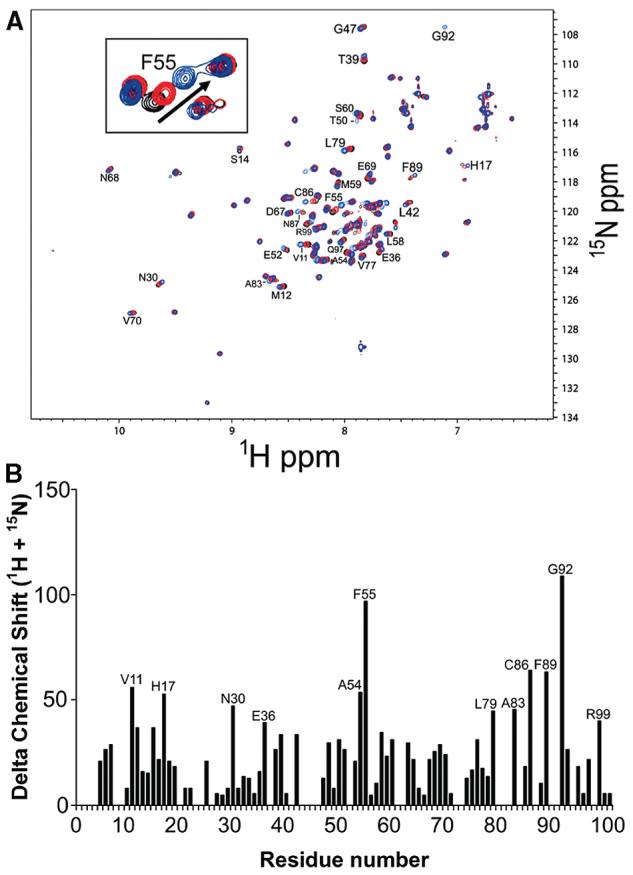
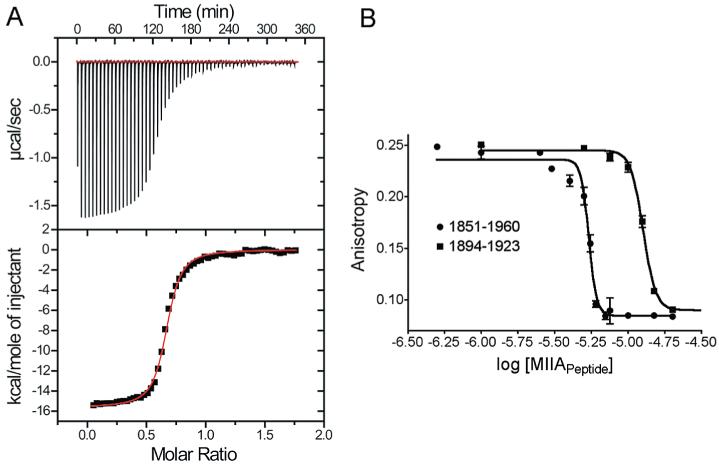
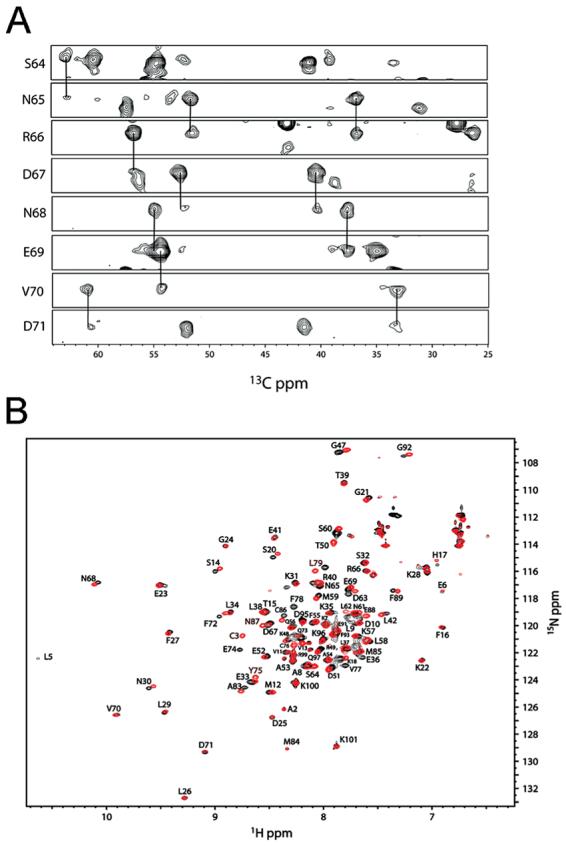
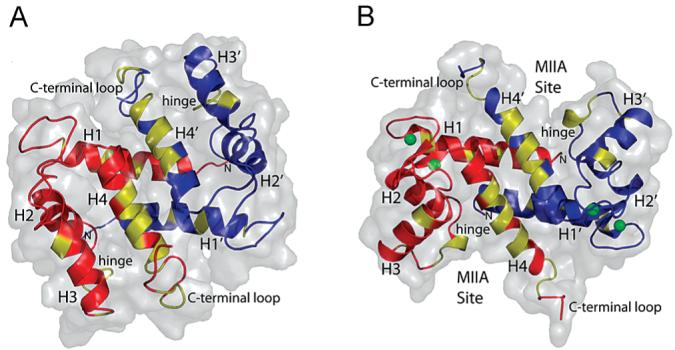
S100A4, also known as mts1, is a member of the S100 family of Ca2+-binding proteins that is directly involved in tumor invasion and metastasis via interactions with specific protein targets, including nonmuscle myosin-IIA (MIIA). Human S100A4 binds two Ca2+ ions with the typical EF-hand exhibiting an affinity that is nearly 1 order of magnitude tighter than that of the pseudo-EF-hand. To examine how Ca2+ modifies the overall organization and structure of the protein, we determined the 1.7 A crystal structure of the human Ca2+-S100A4. Ca2+ binding induces a large reorientation of helix 3 in the typical EF-hand. This reorganization exposes a hydrophobic cleft that is comprised of residues from the hinge region,helix 3, and helix 4, which afford specific target recognition and binding. The Ca2+-dependent conformational change is required for S100A4 to bind peptide sequences derived from the C-terminal portion of the MIIA rod with submicromolar affinity. In addition, the level of binding of Ca2+ to both EF-hands increases by 1 order of magnitude in the presence of MIIA. NMR spectroscopy studies demonstrate that following titration with a MIIA peptide, the largest chemical shift perturbations and exchange broadening effects occur for residues in the hydrophobic pocket of Ca2+-S100A4. Most of these residues are not exposed in apo-S100A4 and explain the Ca2+ dependence of formation of theS100A4-MIIA complex. These studies provide the foundation for understanding S100A4 target recognition and may support the development of reagents that interfere with S100A4 function.
Figures
References
-
- Donato R. S100: A multigenic family of calcium-modulated proteins of the EF-hand type with intracellular and extracellular functional roles. Int. J. Biochem. Cell Biol. 2001;33:637–668. - PubMed
-
- Marenholz I, Heizmann CW, Fritz G. S100 proteins in mouse and man: From evolution to function and pathology (including an update of the nomenclature) Biochem. Biophys. Res. Commun. 2004;322:1111–1122. - PubMed
-
- Zimmer DB, Cornwall EH, Landar A, Song W. The S100 protein family: History, function, and expression. Brain Res. Bull. 1995;37:417–429. - PubMed
-
- Grigorian MS, Tulchinsky EM, Zain S, Ebralidze AK, Kramerov DA, Kriajevska MV, Georgiev GP, Lukanidin EM. The mts1 gene and control of tumor metastasis. Gene. 1993;135:229–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
