

Review
doi: 10.1016/j.devcel.2008.03.015.
Linking kinetochore-microtubule binding to the spindle checkpoint
Affiliations
- PMID: 18410725
- PMCID: PMC2696048
- DOI: 10.1016/j.devcel.2008.03.015
Item in Clipboard
Review
Linking kinetochore-microtubule binding to the spindle checkpoint
Dev Cell.
2008 Apr.
Abstract
The spindle checkpoint blocks cell-cycle progression until chromosomes are properly attached to the mitotic spindle. Popular models propose that checkpoint proteins associate with kinetochores to produce a "wait anaphase" signal that inhibits anaphase. Recent data suggest that a two-state switch results from using the same kinetochore proteins to bind microtubules and checkpoint proteins. At least eight protein kinases are implicated in spindle checkpoint signaling, arguing that a traditional signal transduction cascade is integral to spindle checkpoint signaling.
Figures
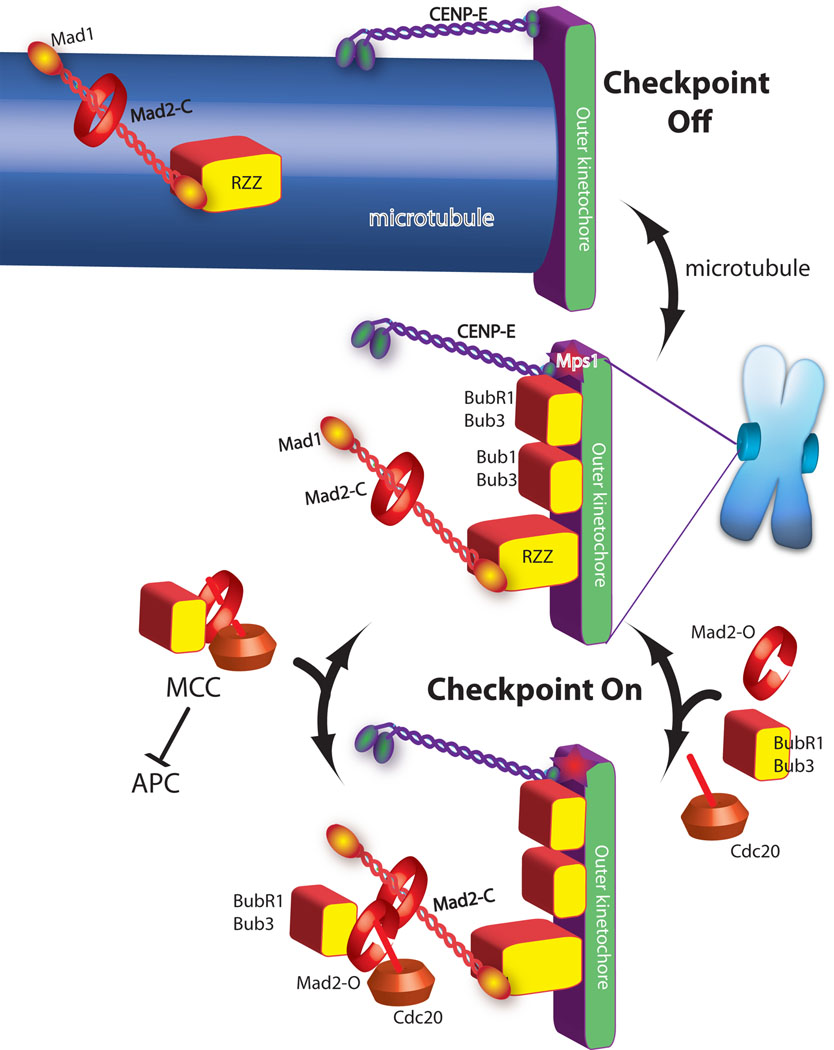
Green and purple are proteins required for microtubule binding. Checkpoint proteins (red and yellow shapes) are recruited to kinetochores that are not binding microtubules (central image). In the absence of microtubules the checkpoint proteins act to catalyze the assembly of the MCC checkpoint complex (bottom image), which diffuses from the kinetochore to inhibit the anaphase promoting complex (APC). Mad2 has both open and closed states and although it is a globular protein it is drawn as a ring to emphasize transitions between these two states. Mad2closed on kinetochores binds Mad2open, which in turn binds Cdc20, Bub1 and Bub3 to form MCC. Microtubule attachment inhibits the signal by two mechanisms. The checkpoint proteins Mad1/Mad2 and RZZ are shown being “stripped” by dynein, which carries them away from the kinetochore by walking towards the minus end of the microtubule (top image). CENP-E activates BubR1 kinase activity unless it binds microtubules, which is also important for silencing the checkpoint.
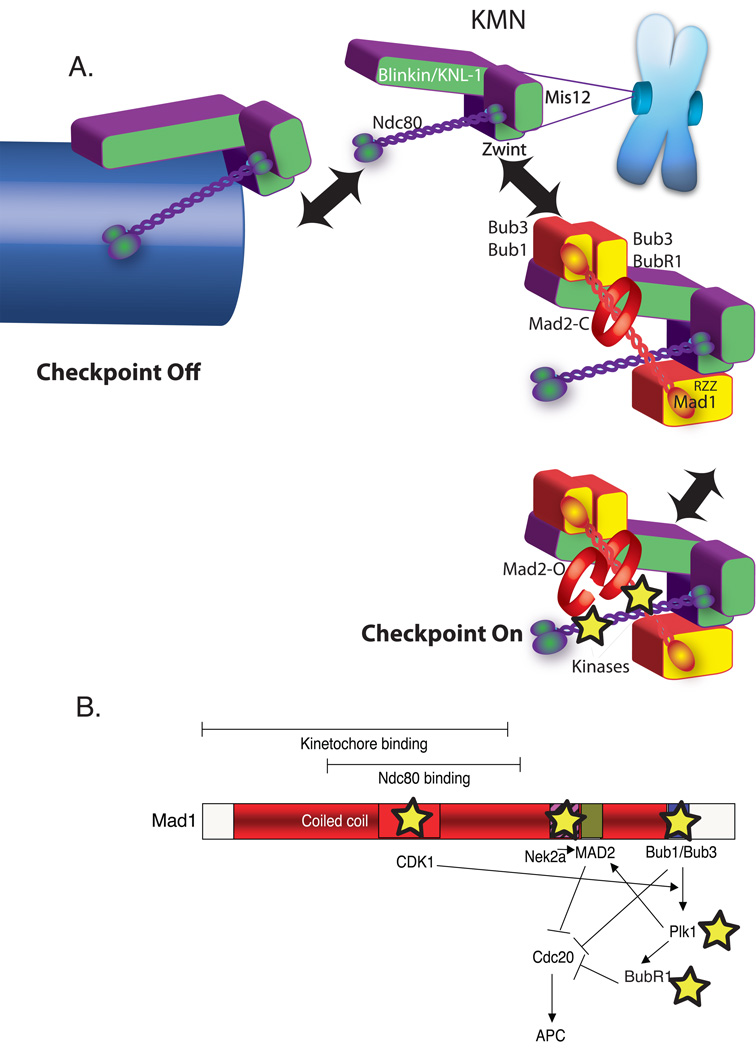
A) Dual role of KMN in the kinetochore as both microtubule anchor and a scaffold to generate spindle checkpoint signals. Overall color scheme is the same as Figure 1. KMN (purple and green) is located in the outer plate of the kinetochore where both binds microtubules (arrow left) and spindle checkpoint protein (arrow right). KMN contains at least two microtubule-binding interfaces one in the KNL-1 subunit and another in the Hec1 subunit of the Ndc80 complex and approximately eight KMNs generate a binding pocket (not shown). In the absence of microtubules KMN has both direct and indirect interactions with checkpoint proteins. KNL-1 binds Bub3/Bub1 and Bub3/BubR1. The Ndc80 subunit can bind a coiled-coil region of Mad1 in a two-hybrid assay. Finally through the Zwint protein, the Mis12 complex binds RZZ, which can strip Mad1 from kinetochores. The kinetochore activates the checkpoint by acting as a scaffold to recruit and activate Bub1 and BubR1 kinases as well as other kinases recently implicated in checkpoint signaling (stars). B) A schematic Map of the Mad1 protein highlighting kinase interactions (stars) and a potential signal transduction network initiated after Mad1 recruitment to the kinetochore.
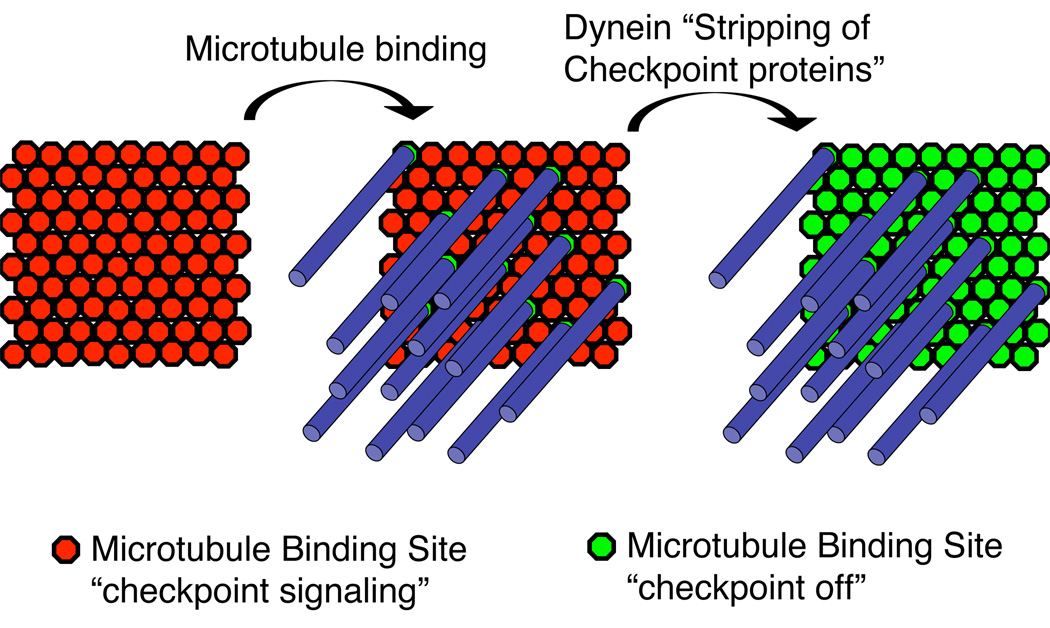
Since there is more KMN than is necessary to bind microtubules there may be two independent steps in checkpoint silencing. First, microtubule binding sterically prevents checkpoint protein binding to potential microtubule binding sites. Second, dynein “strips” checkpoint proteins from surrounding KMN complexes that are not associated with microtubules.
Similar articles
-
Spindle checkpoint proteins and chromosome-microtubule attachment in budding yeast.J Cell Biol. 2004 Feb 16;164(4):535-46. doi: 10.1083/jcb.200308100. Epub 2004 Feb 9. J Cell Biol. 2004. PMID: 14769859 Free PMC article.
-
Bub1 and aurora B cooperate to maintain BubR1-mediated inhibition of APC/CCdc20.J Cell Sci. 2005 Aug 15;118(Pt 16):3639-52. doi: 10.1242/jcs.02487. Epub 2005 Jul 26. J Cell Sci. 2005. PMID: 16046481
-
BubR1 is essential for kinetochore localization of other spindle checkpoint proteins and its phosphorylation requires Mad1.J Cell Biol. 2002 Aug 5;158(3):487-96. doi: 10.1083/jcb.200204048. Epub 2002 Aug 5. J Cell Biol. 2002. PMID: 12163471 Free PMC article.
-
Attachment and tension in the spindle assembly checkpoint.J Cell Sci. 2002 Sep 15;115(Pt 18):3547-55. doi: 10.1242/jcs.00029. J Cell Sci. 2002. PMID: 12186941 Review.
-
How the SAC gets the axe: Integrating kinetochore microtubule attachments with spindle assembly checkpoint signaling.Bioarchitecture. 2015;5(1-2):1-12. doi: 10.1080/19490992.2015.1090669. Epub 2015 Oct 2. Bioarchitecture. 2015. PMID: 26430805 Free PMC article. Review.
Cited by
-
Spindle assembly checkpoint: the third decade.Philos Trans R Soc Lond B Biol Sci. 2011 Dec 27;366(1584):3595-604. doi: 10.1098/rstb.2011.0072. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 22084386 Free PMC article. Review.
-
Bub3p facilitates spindle checkpoint silencing in fission yeast.Mol Biol Cell. 2009 Dec;20(24):5096-105. doi: 10.1091/mbc.e09-09-0762. Mol Biol Cell. 2009. PMID: 19846658 Free PMC article.
-
Kinetochore-microtubule interactions: steps towards bi-orientation.EMBO J. 2010 Dec 15;29(24):4070-82. doi: 10.1038/emboj.2010.294. Epub 2010 Nov 23. EMBO J. 2010. PMID: 21102558 Free PMC article. Review.
-
Connecting the microtubule attachment status of each kinetochore to cell cycle arrest through the spindle assembly checkpoint.Chromosoma. 2015 Dec;124(4):463-80. doi: 10.1007/s00412-015-0515-z. Epub 2015 Apr 28. Chromosoma. 2015. PMID: 25917595 Review.
-
Nearly complete deletion of BubR1 causes microcephaly through shortened mitosis and massive cell death.Hum Mol Genet. 2019 Jun 1;28(11):1822-1836. doi: 10.1093/hmg/ddz022. Hum Mol Genet. 2019. PMID: 30668728 Free PMC article.
References
-
- Basto R, Gomes R, Karess RE. Rough deal and Zw10 are required for the metaphase checkpoint in Drosophila. Nature Cell Biology. 2000;2:939–943. - PubMed
-
- Baumann C, Korner R, Hofmann K, Nigg EA. PICH, a centromere-associated SNF2 family ATPase, is regulated by Plk1 and required for the spindle checkpoint. Cell. 2007;128:101–114. - PubMed
-
- Brady DM, Hardwick KG. Complex formation between Mad1p, Bub1p and Bub3p is crucial for spindle checkpoint function. Curr. Biol. 2000;10:675–678. - PubMed
-
- Chan GK, Jablonski SA, Starr DA, Goldberg ML, Yen TJ. Human Zw10 and ROD are mitotic checkpoint proteins that bind to kinetochores. Nat Cell Biol. 2000;2:944–947. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
