

Organellar dynamics during the cell cycle of Toxoplasma gondii
- PMID: 18411248
- PMCID: PMC6810632
- DOI: 10.1242/jcs.021089
Organellar dynamics during the cell cycle of Toxoplasma gondii
Abstract
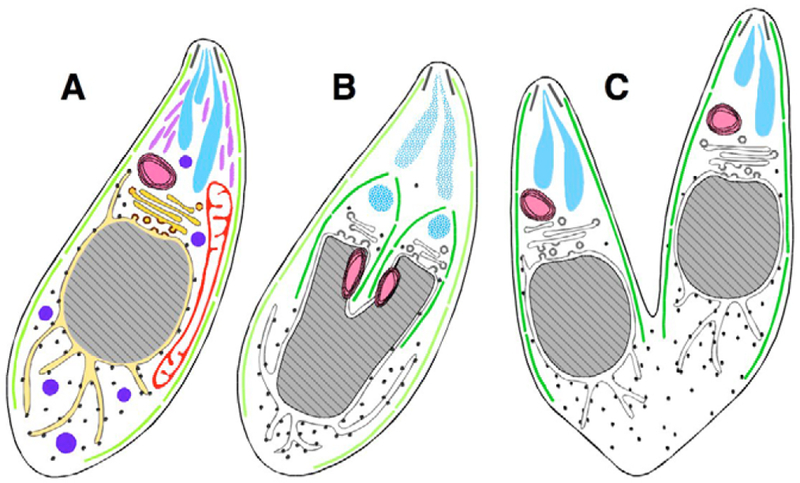
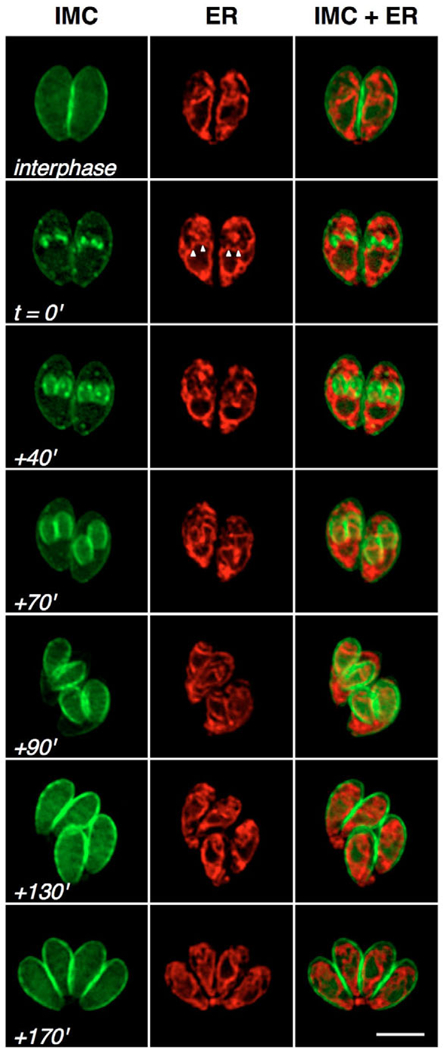
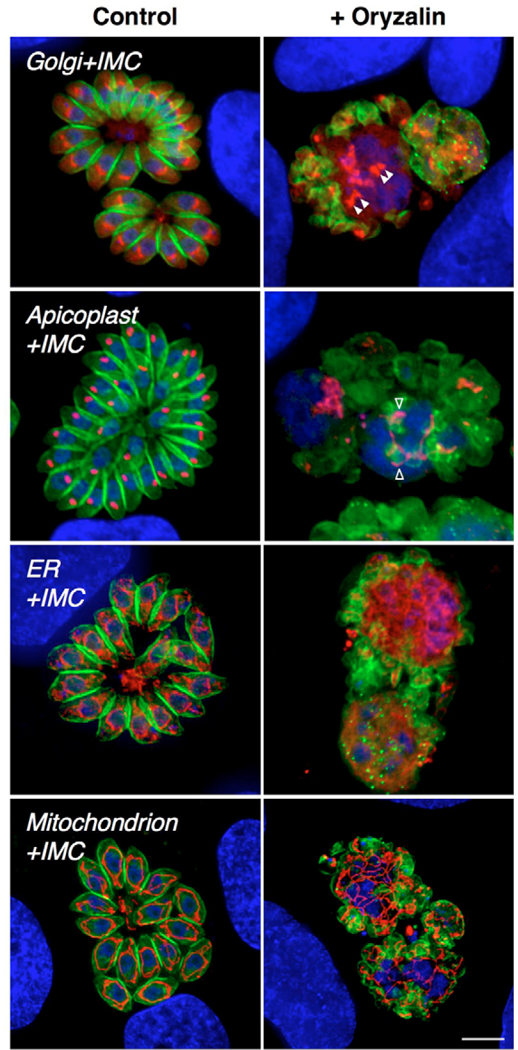
The protozoan phylum Apicomplexa encompasses approximately 5000 species of obligate intracellular parasites, including those responsible for malaria and toxoplasmosis. Rather than dividing by binary fission, apicomplexans use a remarkable mechanism for replication, assembling daughters de novo within the cytoplasm. Here, we exploit time-lapse microscopy of fluorescent markers targeted to various subcellular structures in Toxoplasma gondii tachyzoites to determine how these unicellular eukaryotes efficiently package a complete set of organelles, maintaining the highly polarized organization necessary for host cell invasion and pathogenesis. Golgi division and elongation of the apicoplast are among the first morphologically observable events, associated with an unusual pattern of centriolar migration. Daughter parasites are assembled on cytoskeletal scaffolding, whose growth proceeds from the apical end, first encapsulating the divided Golgi. Further extension of the cytoskeletal scaffold results in partitioning of the apicoplast, nucleus, endoplasmic reticulum, and finally the mitochondrion, which enters the developing daughters rapidly, but only very late during the division cycle. The specialized secretory organelles (micronemes and rhoptries) form de novo. This distinctive pattern of replication -- in which organellar segregation spans approximately 75% of the cell cycle, completely encompassing S phase -- suggests an unusual mechanism of cell cycle regulation.
Figures




References
-
- Aikawa M (1971). Parasitological review. Plasmodium: the fine structure of malarial parasites. Exp. Parasitol 30, 284–320. - PubMed
-
- Bannister LH, Hopkins JM, Fowler RE, Krishna S and Mitchell GH (2000a). A brief illustrated guide to the ultrastructure of Plasmodium falciparum asexual blood stages. Parasitol. Today 16, 427–433. - PubMed
-
- Bannister LH, Hopkins JM, Fowler RE, Krishna S and Mitchell GH (2000b).Ultrastructure of rhoptry development in Plasmodium falciparum erythrocytic schizonts. Parasitology 121, 273–287. - PubMed
-
- Bereiter-Hahn J and Voth M (1994). Dynamics of mitochondria in living cells: shape changes, dislocations, fusion, and fission of mitochondria. Microsc. Res. Tech 27, 198–219. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
