

Spinophilin participates in information transfer at immunological synapses
- PMID: 18411312
- PMCID: PMC2315669
- DOI: 10.1083/jcb.200711149
Spinophilin participates in information transfer at immunological synapses
Abstract
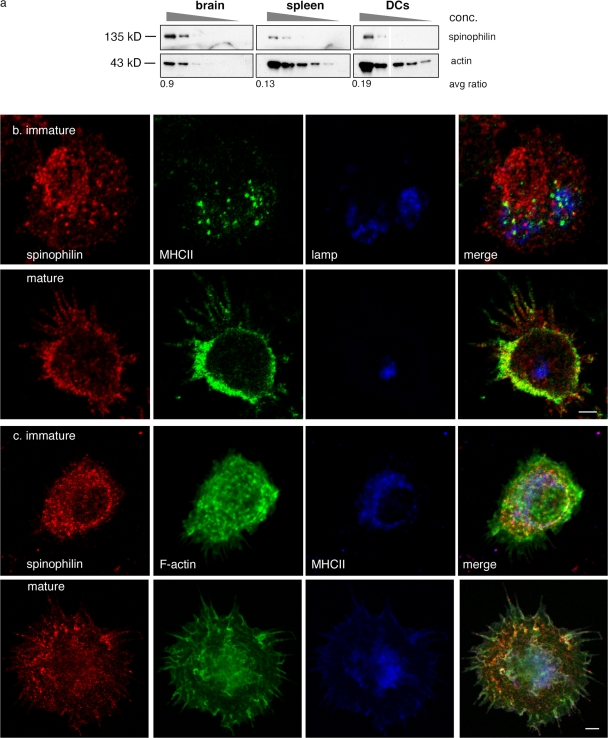
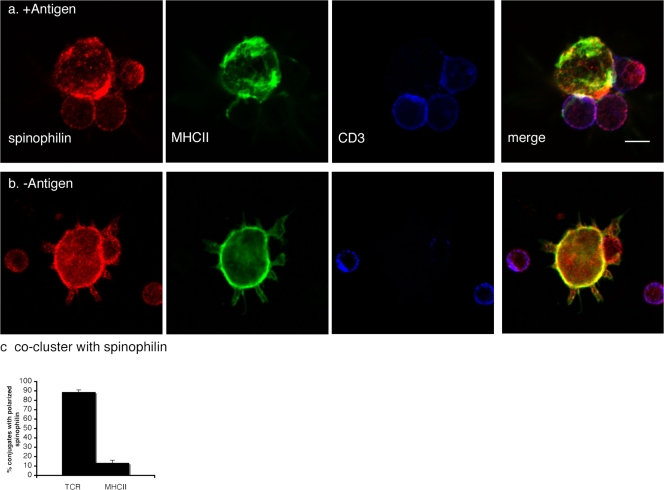
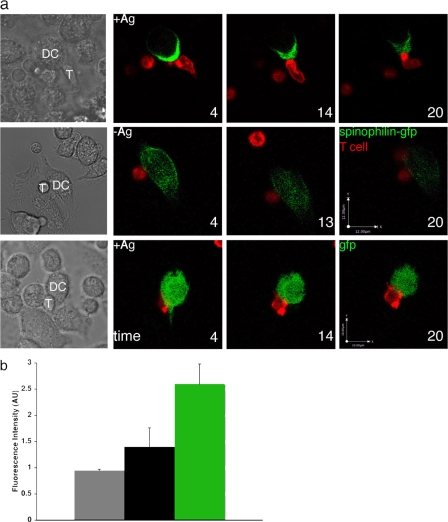
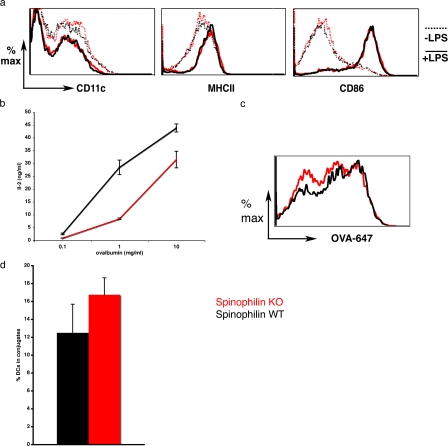
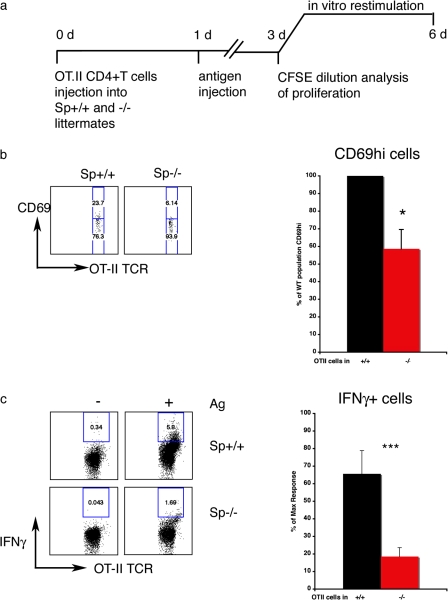
The adaptive immune response is initiated by the presentation of peptides bound to major histocompatibility complex molecules on dendritic cells (DCs) to antigen-specific T lymphocytes at a junction termed the immunological synapse. Although much attention has been paid to cytoplasmic events on the T cell side of the synapse, little is known concerning events on the DC side. We have sought signal transduction components of the neuronal synapse that were also expressed by DCs. One such protein is spinophilin, a scaffolding protein of neuronal dendritic spines that regulates synaptic transmission. In inactive, immature DCs, spinophilin is located throughout the cytoplasm but redistributes to the plasma membrane upon stimulus-induced maturation. In DCs interacting with T cells, spinophilin is polarized dynamically to contact sites in an antigen-dependent manner. It is also required for optimal T cell activation because DCs derived from mice lacking spinophilin exhibit defects in antigen presentation both in vitro and in vivo. Thus, spinophilin may play analogous roles in information transfer at both neuronal and immunological synapses.
Figures
References
-
- Allen, P.B. 2004. Functional plasticity in the organization of signaling complexes in the striatum. Parkinsonism Relat. Disord. 10:287–292. - PubMed
-
- Allen, P.B., V. Zachariou, P. Svenningsson, A.C. Lepore, D. Centonze, C. Costa, S. Rossi, G. Bender, G. Chen, J. Feng, et al. 2006. Distinct roles for spinophilin and neurabin in dopamine-mediated plasticity. Neuroscience. 140:897–911. - PubMed
-
- Benvenuti, F., S. Hugues, M. Walmsley, S. Ruf, L. Fetler, M. Popoff, V.L. Tybulewicz, and S. Amigorena. 2004. Requirement of Rac1 and Rac2 expression by mature dendritic cells for T cell priming. Science. 305:1150–1153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
