

Aldosterone and the autocrine modulation of potassium currents and oxidative stress in the diabetic rat heart
- PMID: 18414392
- PMCID: PMC2439528
- DOI: 10.1038/bjp.2008.114
Aldosterone and the autocrine modulation of potassium currents and oxidative stress in the diabetic rat heart
Abstract
Background and purpose: Aldosterone plays a major role in cardiac pathology. This study was designed to investigate the role of cardiac aldosterone in modulating K(+) currents and oxidative stress in the streptozotocin-induced diabetic rat heart.
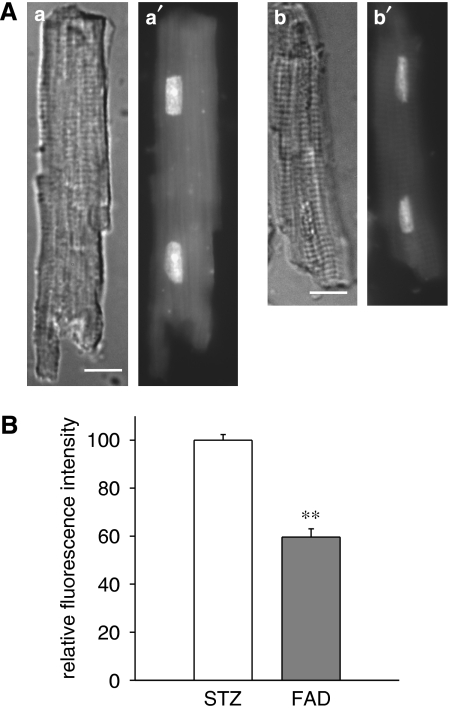
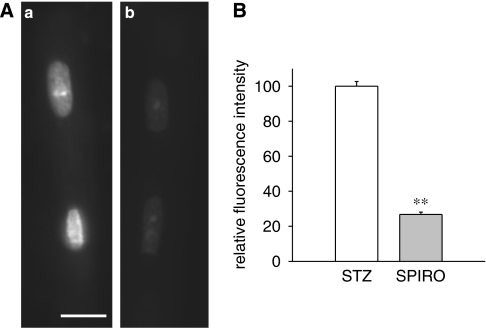
Experimental approach: Transient and sustained K(+) currents were measured in ventricular myocytes by voltage clamp. Plasma and cellular aldosterone were measured by ELISA. Fluorescent dihydroethidium (DHE) was used to assess superoxide ions as markers of oxidative stress.
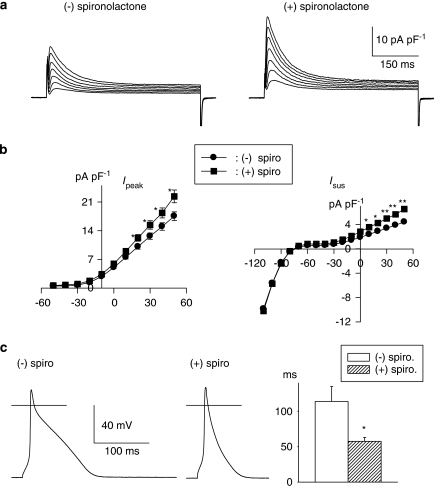
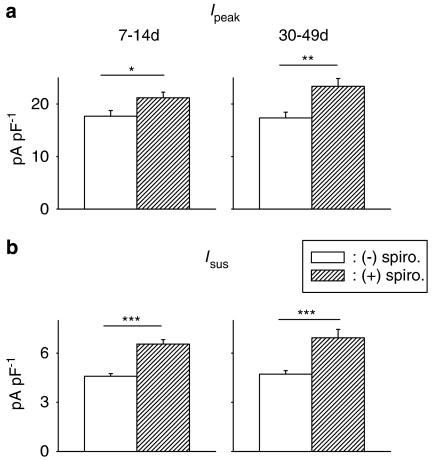
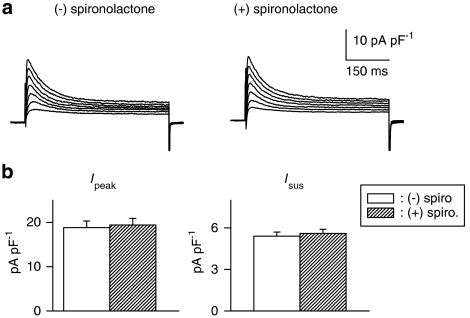
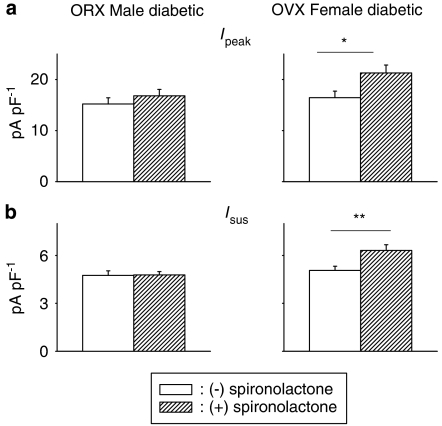
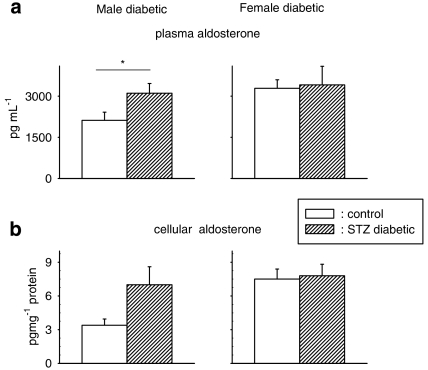
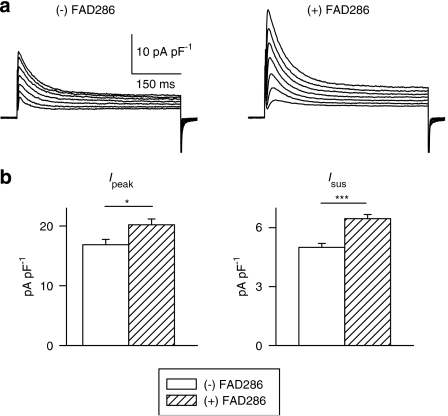
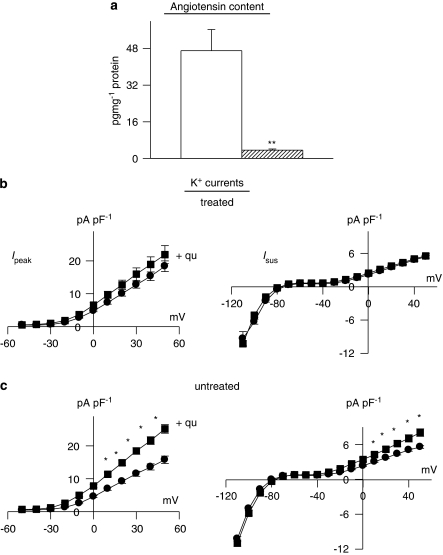
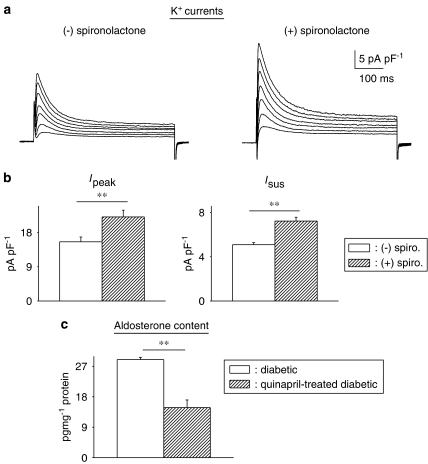
Key results: The mineralocorticoid antagonist spironolactone (1 microM, 5-9 h) significantly augmented both K(+) currents in diabetic males, with a concomitant shortening of the action potential but had no effect in myocytes from control males or from diabetic females. Effects of spironolactone were restored in ovariectomized diabetic females and abolished in orchidectomized diabetic males. The aldosterone synthase inhibitor FAD286 (1 microM, 5-9 h) significantly augmented K(+) currents in cells from diabetic males, but not females. Spironolactone and FAD286 significantly reduced oxidative stress in cells from diabetic males. Plasma aldosterone content was elevated in diabetic males (relative to control), but not in females. Cellular aldosterone was also elevated, but not significantly. The elevation in aldosterone was only partly dependent on a concomitant increase in cellular angiotensin II.
Conclusions and implications: A gender-related, sex-hormone-dependent elevation in plasma and cardiac cell aldosterone contributed to oxidative stress and to attenuation of K(+) currents in diabetic male rats. Aldosterone may thus contribute to diabetes-associated cardiac arrhythmias. Aldosterone elevation was partly related to levels of angiotensin II, but residual, angiotensin II-independent, aldosterone maintains functional relevance.
Figures
References
-
- Abo K, Ishida Y, Yoshida R, Hozumi T, Ueno H, Shiotani H, et al. Torsade de Pointes in NIDDM with long QT intervals. Diabetes Care. 1996;9:1010. - PubMed
-
- Bell DSH. Diabetic cardiomyopathy. Diabetes Care. 2003;26:2949–2951. - PubMed
-
- Bos R, Mougenot N, Findji L, Mediani O, Vanhoutte PM, Lechat P. Inhibition of catecholamine-induced cardiac fibrosis by an aldosterone antagonist. J Cardiovasc Pharmacol. 2005;45:8–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
