

Cyclic and sleep-like spontaneous alternations of brain state under urethane anaesthesia
- PMID: 18414674
- PMCID: PMC2289875
- DOI: 10.1371/journal.pone.0002004
Cyclic and sleep-like spontaneous alternations of brain state under urethane anaesthesia
Abstract
Background: Although the induction of behavioural unconsciousness during sleep and general anaesthesia has been shown to involve overlapping brain mechanisms, sleep involves cyclic fluctuations between different brain states known as active (paradoxical or rapid eye movement: REM) and quiet (slow-wave or non-REM: nREM) stages whereas commonly used general anaesthetics induce a unitary slow-wave brain state.
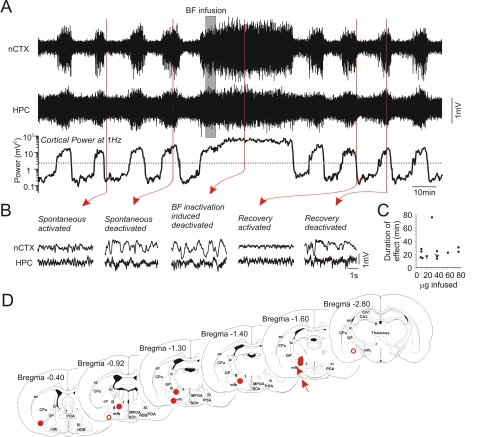
Methodology/principal findings: Long-duration, multi-site forebrain field recordings were performed in urethane-anaesthetized rats. A spontaneous and rhythmic alternation of brain state between activated and deactivated electroencephalographic (EEG) patterns was observed. Individual states and their transitions resembled the REM/nREM cycle of natural sleep in their EEG components, evolution, and time frame ( approximately 11 minute period). Other physiological variables such as muscular tone, respiration rate, and cardiac frequency also covaried with forebrain state in a manner identical to sleep. The brain mechanisms of state alternations under urethane also closely overlapped those of natural sleep in their sensitivity to cholinergic pharmacological agents and dependence upon activity in the basal forebrain nuclei that are the major source of forebrain acetylcholine. Lastly, stimulation of brainstem regions thought to pace state alternations in sleep transiently disrupted state alternations under urethane.
Conclusions/significance: Our results suggest that urethane promotes a condition of behavioural unconsciousness that closely mimics the full spectrum of natural sleep. The use of urethane anaesthesia as a model system will facilitate mechanistic studies into sleep-like brain states and their alternations. In addition, it could also be exploited as a tool for the discovery of new molecular targets that are designed to promote sleep without compromising state alternations.
Conflict of interest statement
Figures










References
-
- Rechtschaffen A, Bergmann BM, Everson CA, Kushida CA, Gilliland MA. Sleep deprivation in the rat: X. Integration and discussion of the findings. 1989. Sleep. 2002;25:68–87. - PubMed
-
- Rechtschaffen A, Bergmann BM. Sleep deprivation in the rat: an update of the 1989 paper. Sleep. 2002;25:18–24. - PubMed
-
- Ocampo_Garces A, Molina E, Rodriguez A, Vivaldi EA. Homeostasis of REM sleep after total and selective sleep deprivation in the rat. J Neurophysiol. 2000;84:2699–2702. - PubMed
-
- Rechtschaffen A, Bergmann BM, Gilliland MA, Bauer K. Effects of method, duration, and sleep stage on rebounds from sleep deprivation in the rat. Sleep. 1999;22:11–31. - PubMed
-
- Ferrara M, De_Gennaro L, Bertini M. Selective slow-wave sleep (SWS) deprivation and SWS rebound: do we need a fixed SWS amount per night? Sleep Res Online. 1999;2:15–19. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
