

The Retinoic Acid Receptor-alpha mediates human T-cell activation and Th2 cytokine and chemokine production
- PMID: 18416830
- PMCID: PMC2394516
- DOI: 10.1186/1471-2172-9-16
The Retinoic Acid Receptor-alpha mediates human T-cell activation and Th2 cytokine and chemokine production
Abstract
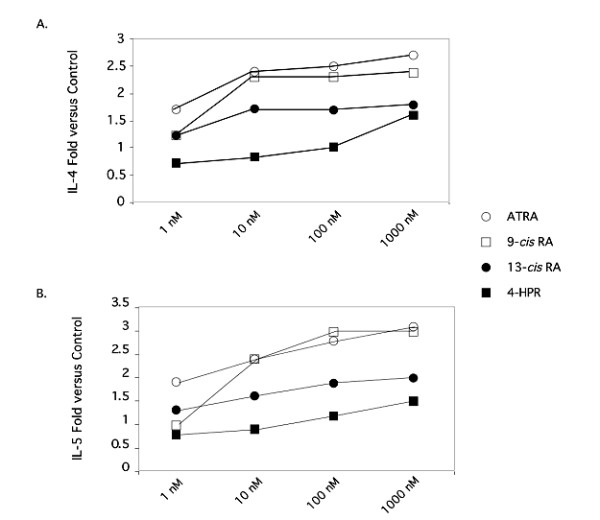
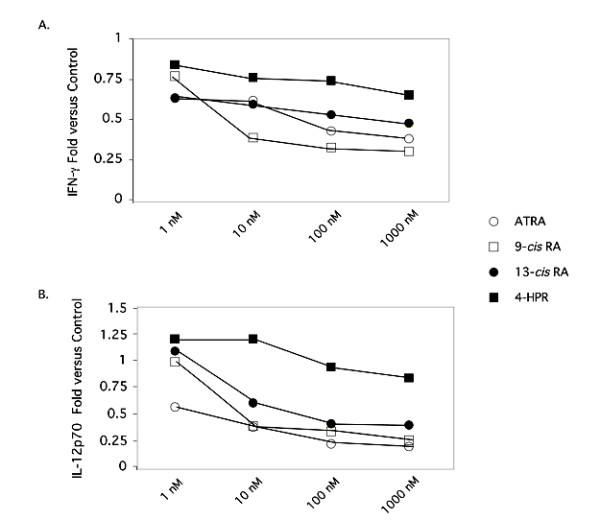
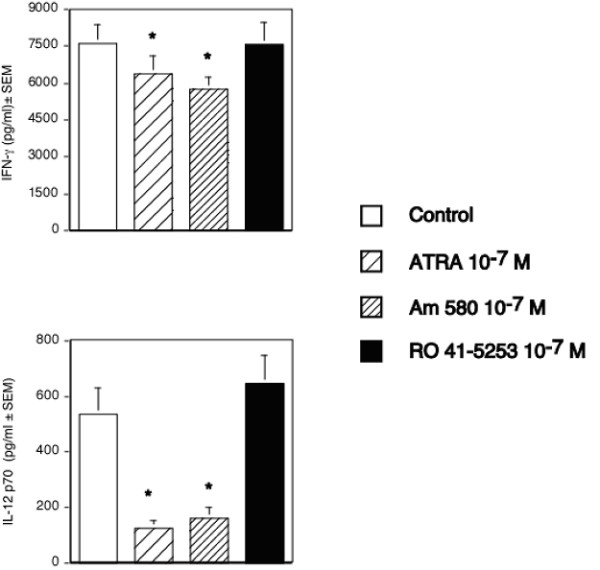
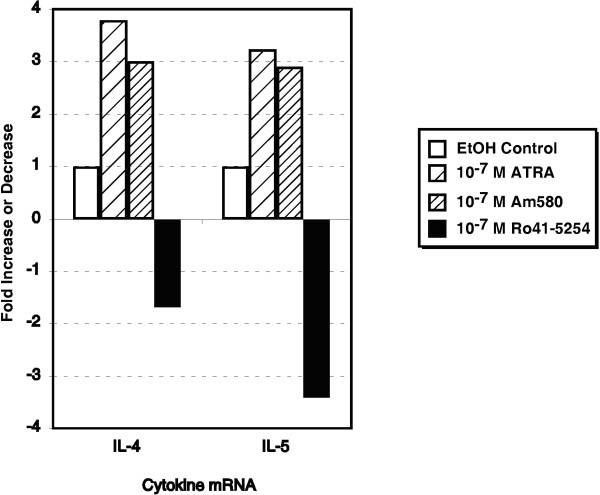
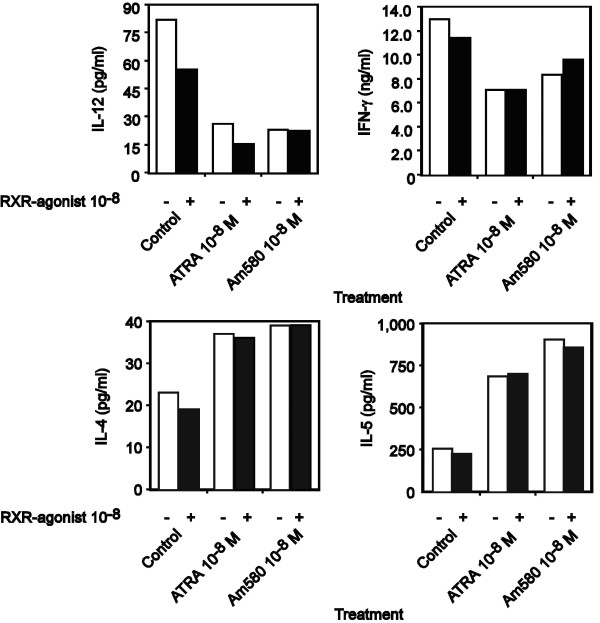
Background: We have recently demonstrated that all-trans-retinoic acid (ATRA) and 9-cis-retinoic acid (9-cis RA) promote IL-4, IL-5 and IL-13 synthesis, while decreasing IFN-gamma and TNF-alpha expression by activated human T cells and reduces the synthesis of IL-12p70 from accessory cells. Here, we have demonstrated that the observed effects using ATRA and 9-cis RA are shared with the clinically useful RAR ligand, 13-cis retinoic acid (13-cis RA), and the retinoic acid receptor-alpha (RAR-alpha)-selective agonist, AM580 but not with the RAR-beta/gamma ligand, 4-hydroxyphenylretinamide (4-HPR).
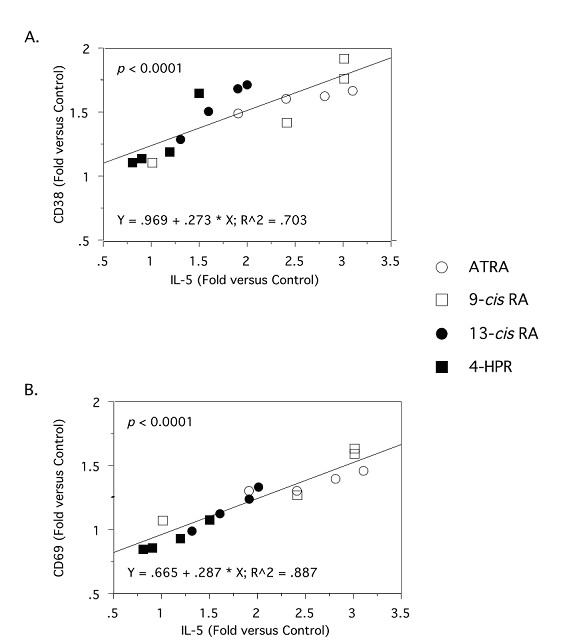
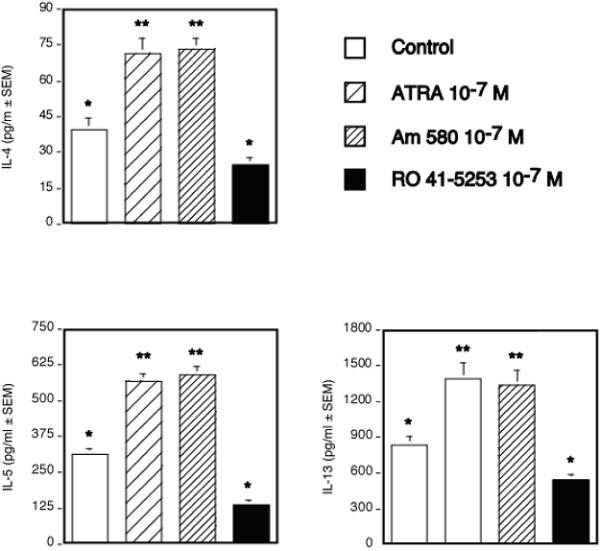
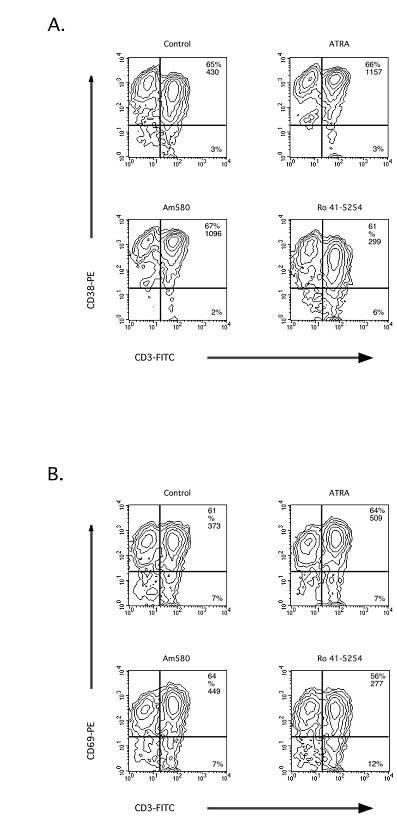
Results: The increase in type 2 cytokine production by these retinoids correlated with the expression of the T cell activation markers, CD69 and CD38. The RAR-alpha-selective agonist, AM580 recapitulated all of the T cell activation and type 2 cytokine-inducing effects of ATRA and 9-cis-RA, while the RAR-alpha-selective antagonist, RO 41-5253, inhibited these effects.
Conclusion: These results strongly support a role for RAR-alpha engagement in the regulation of genes and proteins involved with human T cell activation and type 2 cytokine production.
Figures
Similar articles
-
The prevention of adipose differentiation of 3T3-L1 cells caused by retinoic acid is elicited through retinoic acid receptor alpha.Life Sci. 1994;55(16):PL307-12. doi: 10.1016/0024-3205(94)90073-6. Life Sci. 1994. PMID: 7934625
-
AM580, a stable benzoic derivative of retinoic acid, has powerful and selective cyto-differentiating effects on acute promyelocytic leukemia cells.Blood. 1996 Feb 15;87(4):1520-31. Blood. 1996. PMID: 8608243
-
Retinoic acid receptor alpha expression correlates with retinoid-induced growth inhibition of human breast cancer cells regardless of estrogen receptor status.Cancer Res. 1997 Jul 1;57(13):2642-50. Cancer Res. 1997. PMID: 9205071
-
Disparate roles of retinoid acid signaling molecules in kidney disease.Am J Physiol Renal Physiol. 2021 May 1;320(5):F683-F692. doi: 10.1152/ajprenal.00045.2021. Epub 2021 Mar 1. Am J Physiol Renal Physiol. 2021. PMID: 33645319 Free PMC article. Review.
-
Regulation of CD8(+) T cell functions by RARgamma.Semin Immunol. 2009 Feb;21(1):2-7. doi: 10.1016/j.smim.2008.07.002. Epub 2008 Aug 20. Semin Immunol. 2009. PMID: 18715802 Free PMC article. Review.
Cited by
-
Universal allogeneic CAR T cells engineered with Sleeping Beauty transposons and CRISPR-CAS9 for cancer immunotherapy.Mol Ther. 2022 Oct 5;30(10):3155-3175. doi: 10.1016/j.ymthe.2022.06.006. Epub 2022 Jun 16. Mol Ther. 2022. PMID: 35711141 Free PMC article.
-
The retinoic acid receptor-α modulators ATRA and Ro415253 reciprocally regulate human IL-5+ Th2 cell proliferation and cytokine expression.Clin Mol Allergy. 2013 Dec 6;11(1):4. doi: 10.1186/1476-7961-11-4. Clin Mol Allergy. 2013. PMID: 24314292 Free PMC article.
-
The effects of all-trans retinoic acid on immune cells and its formulation design for vaccines.AAPS J. 2021 Feb 24;23(2):32. doi: 10.1208/s12248-021-00565-1. AAPS J. 2021. PMID: 33629139 Review.
-
Rapid profiling of transcription factor-cofactor interaction networks reveals principles of epigenetic regulation.Nucleic Acids Res. 2024 Sep 23;52(17):10276-10296. doi: 10.1093/nar/gkae706. Nucleic Acids Res. 2024. PMID: 39166482 Free PMC article.
-
ATRA mechanically reprograms pancreatic stellate cells to suppress matrix remodelling and inhibit cancer cell invasion.Nat Commun. 2016 Sep 7;7:12630. doi: 10.1038/ncomms12630. Nat Commun. 2016. PMID: 27600527 Free PMC article.
References
-
- Napoli JL. Quantification of physiological levels of retinoic acid. Methods in Enzymology. 1986;123:112–124. - PubMed
-
- Kojima R, Fujimori T, Kiyota N, Toriya Y, Fukuda T, Ohashi T, Sato T, Yoshizawa Y, Takeyama K, Mani H, Masushige D, Kato S. In vivo isomerization of retinoic acids. Rapid isomer exchange and gene expression. Journal of Biological Chemistry. 1994;269:32700–32707. - PubMed
-
- Ziouzenkova O, Orashanu G, Sukhova G, Lau E, Berger JP, Tang G, Krinsky NI, Dolnikowski GG, Plutzky J. Asymmetric cleavage of {beta} -carotene yields a transcriptional repressor of RXR and PPAR responses. Mol Endocrinol. 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
