

Unique Asn-linked oligosaccharides of the human pathogen Entamoeba histolytica
- PMID: 18417475
- PMCID: PMC2440624
- DOI: 10.1074/jbc.M800725200
Unique Asn-linked oligosaccharides of the human pathogen Entamoeba histolytica
Abstract
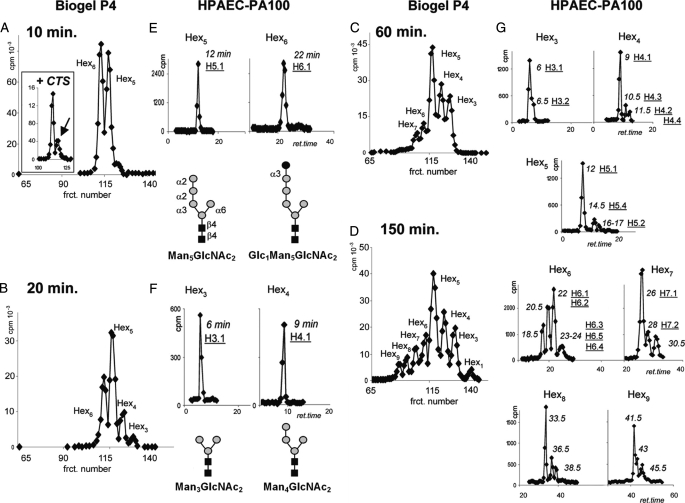
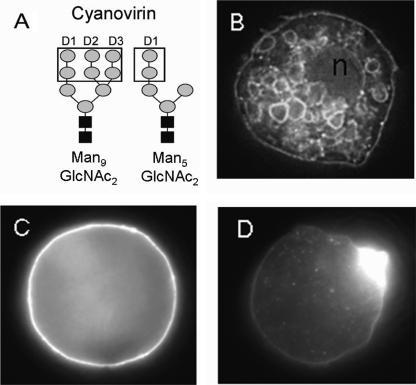
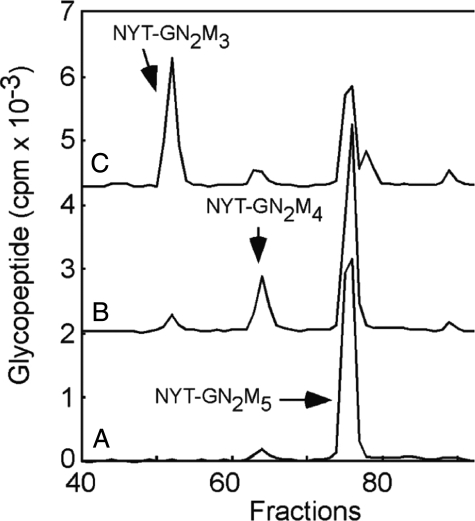
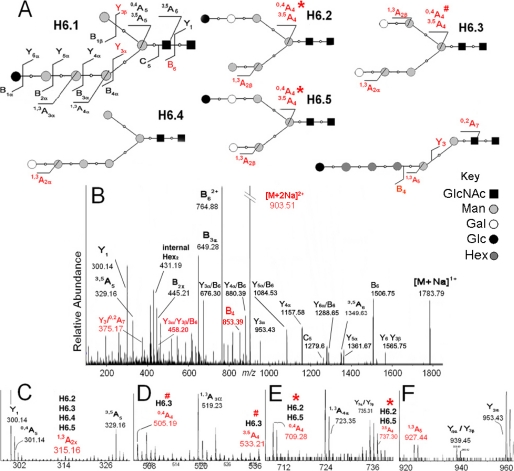
N-Glycans of Entamoeba histolytica, the protist that causes amebic dysentery and liver abscess, are of great interest for multiple reasons. E. histolytica makes an unusual truncated N-glycan precursor (Man(5)GlcNAc(2)), has few nucleotide sugar transporters, and has a surface that is capped by the lectin concanavalin A. Here, biochemical and mass spectrometric methods were used to examine N-glycan biosynthesis and the final N-glycans of E. histolytica with the following conclusions. Unprocessed Man(5)GlcNAc(2), which is the most abundant E. histolytica N-glycan, is aggregated into caps on the surface of E. histolytica by the N-glycan-specific, anti-retroviral lectin cyanovirin-N. Glc(1)Man(5)GlcNAc(2), which is made by a UDP-Glc: glycoprotein glucosyltransferase that is part of a conserved N-glycan-dependent endoplasmic reticulum quality control system for protein folding, is also present in mature N-glycans. A swainsonine-sensitive alpha-mannosidase trims some N-glycans to biantennary Man(3)GlcNAc(2). Complex N-glycans of E. histolytica are made by the addition of alpha1,2-linked Gal to both arms of small oligomannose glycans, and Gal residues are capped by one or more Glc. In summary, E. histolytica N-glycans include unprocessed Man(5)GlcNAc(2), which is a target for cyanovirin-N, as well as unique, complex N-glycans containing Gal and Glc.
Figures





References
-
- Haque, R., Huston, C. D., Hughes, M., Houpt, E., and Petri, W. A., Jr. (2003) N. Engl. J. Med. 348 1565-1573 - PubMed
-
- Stanley, S. L. (2003) Lancet 361 1025-1034 - PubMed
-
- Helenius, A., and Aebi, M. (2004) Annu. Rev. Biochem. 73 1019-1049 - PubMed
-
- Trombetta, E. S., and Parodi, A. J. (2003) Annu. Rev. Cell Dev. Biol. 19 649-676 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
