

Basolateral entry and release of New and Old World arenaviruses from human airway epithelia
- PMID: 18417570
- PMCID: PMC2395129
- DOI: 10.1128/JVI.00100-08
Basolateral entry and release of New and Old World arenaviruses from human airway epithelia
Abstract
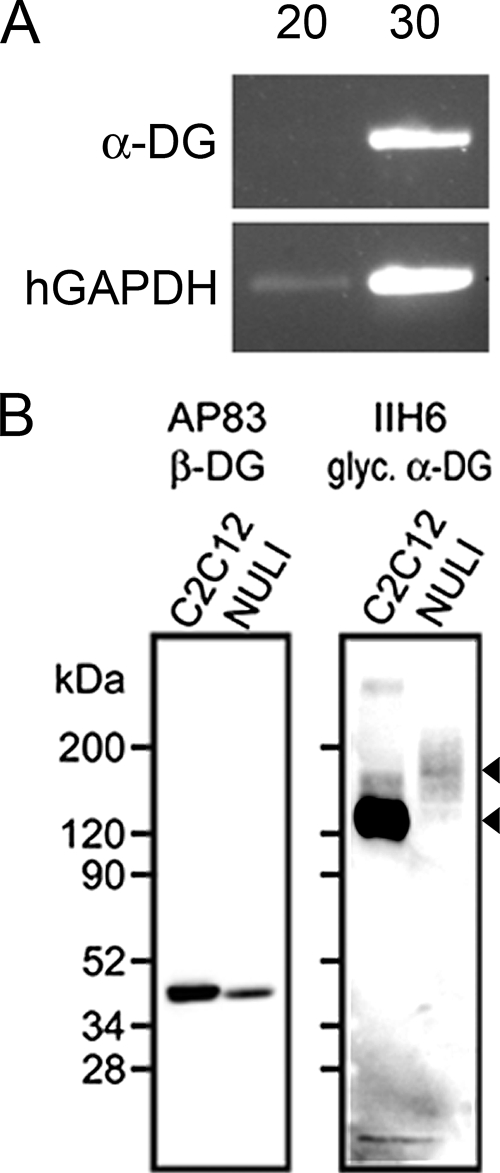
Transmission of arenaviruses from rodent hosts to humans is generally thought to occur through inhalation or ingestion of dust or droplets containing viral particles. Here we demonstrate that two identified arenavirus receptors, alpha-dystroglycan (alpha-DG) and transferrin receptor 1 (TfR1), are expressed in polarized human airway epithelia. Lymphocytic choriomeningitis virus strains with high or low alpha-DG affinity and Junin virus, which binds TfR1, efficiently infected polarized epithelia only when applied to the basolateral surface or when injury compromised tight junction integrity. Viral egress from infected epithelia exhibited basolateral polarity. This study demonstrates that respiratory entry of arenaviruses occurs via basolateral receptors.
Figures
References
-
- Baldridge, J. R., B. D. Pearce, B. S. Parekh, and M. J. Buchmeier. 1993. Teratogenic effects of neonatal arenavirus infection on the developing rat cerebellum are abrogated by passive immunotherapy. Virology 197669-677. - PubMed
-
- Buchmeier, M. J., M. D. Bowen, and C. J. Peters. 2001. Arenaviridae: the viruses and their replication, p. 1635-1668. In D. M. Knipe and P. M. Howley (ed.), Fields virology. Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Cao, W., M. D. Henry, P. Borrow, H. Yamada, J. H. Elder, E. V. Ravkov, S. T. Nichol, R. W. Compans, K. P. Campbell, and M. B. Oldstone. 1998. Identification of alpha-dystroglycan as a receptor for lymphocytic choriomeningitis virus and Lassa fever virus. Science 2822079-2081. - PubMed
-
- Chu, J. J., and M. L. Ng. 2002. Infection of polarized epithelial cells with flavivirus West Nile: polarized entry and egress of virus occur through the apical surface. J. Gen. Virol. 832427-2435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
