

Analysis of DBC1 and its homologs suggests a potential mechanism for regulation of sirtuin domain deacetylases by NAD metabolites
- PMID: 18418069
- PMCID: PMC2423810
- DOI: 10.4161/cc.7.10.5883
Analysis of DBC1 and its homologs suggests a potential mechanism for regulation of sirtuin domain deacetylases by NAD metabolites
Abstract
Deleted in Breast Cancer-1 (DBC1) and its paralog CARP-1 are large multi-domain proteins, with a nuclear or perinuclear localization, and a role in promoting apoptosis upon processing by caspases. Recent studies on human DBC1 show that it is a specific inhibitor of the sirtuin-type deacetylase, Sirt1, which deacetylates histones and p53. Using sensitive sequence profile searches and HMM-HMM comparisons we show that the central conserved globular domain present in the DBC1 and it homologs from diverse eukaryotes is a catalytically inactive version of the Nudix hydrolase (MutT) domain. Given that Nudix domains are known to bind nucleoside diphosphate sugars and NAD, we predict that this domain in DBC1 and its homologs binds NAD metabolites such as ADP-ribose. Hence, we propose that DBC1 and its homologs are likely to regulate the activity of SIRT1 or related deacetylases by sensing the soluble products or substrates of the NAD-dependent deacetylation reaction. The complex domain architectures of the members of the DBC1 family, which include fusions to the RNA-binding S1-like domain, the DNA-binding SAP domain and EF-hand domains, suggest that they are likely to function as integrators of distinct regulatory signals including chromatin protein modification, soluble compounds in NAD metabolism, apoptotic stimuli and RNA recognition.
Figures
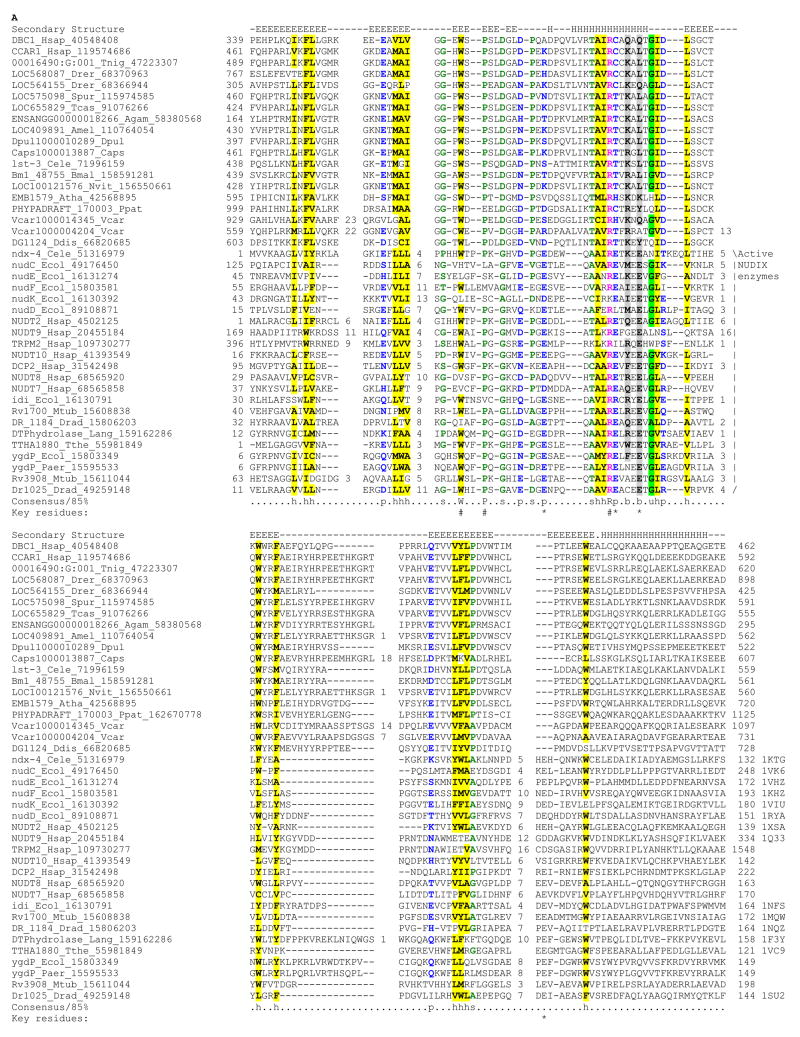

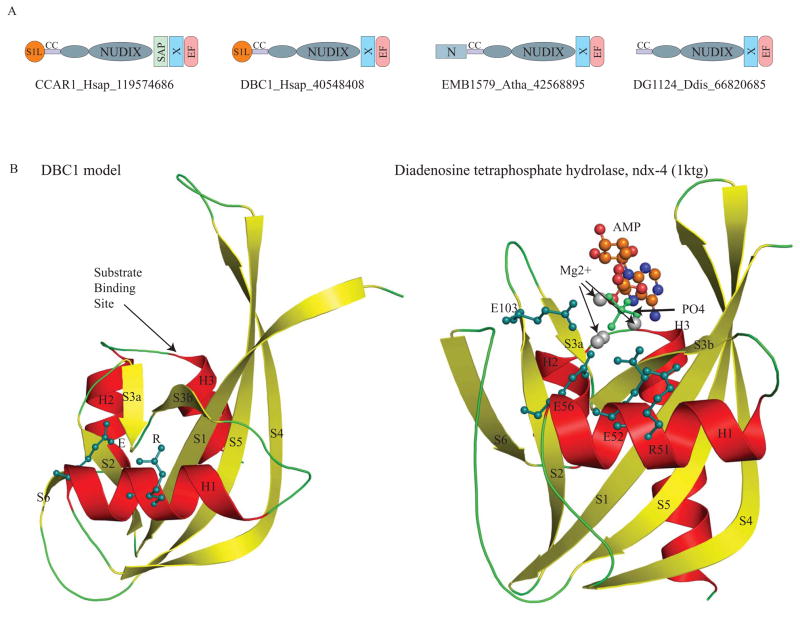
References
-
- Sundararajan R, Chen G, Mukherjee C, White E. Caspase-dependent processing activates the proapoptotic activity of deleted in breast cancer-1 during tumor necrosis factor-alpha-mediated death signaling. Oncogene. 2005;24:4908–20. - PubMed
-
- Rishi AK, Zhang L, Yu Y, Jiang Y, Nautiyal J, Wali A, Fontana JA, Levi E, Majumdar AP. Cell cycle- and apoptosis-regulatory protein-1 is involved in apoptosis signaling by epidermal growth factor receptor. J Biol Chem. 2006;281:13188–98. - PubMed
-
- Kim JE, Chen J, Lou Z. DBC1 is a negative regulator of SIRT1. Nature. 2008;451:583–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous