

Topoisomerase IIIalpha is required for normal proliferation and telomere stability in alternative lengthening of telomeres
- PMID: 18418389
- PMCID: PMC2396391
- DOI: 10.1038/emboj.2008.74
Topoisomerase IIIalpha is required for normal proliferation and telomere stability in alternative lengthening of telomeres
Abstract
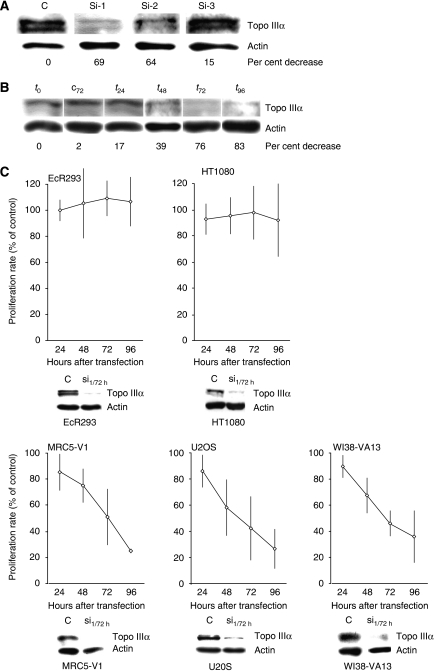
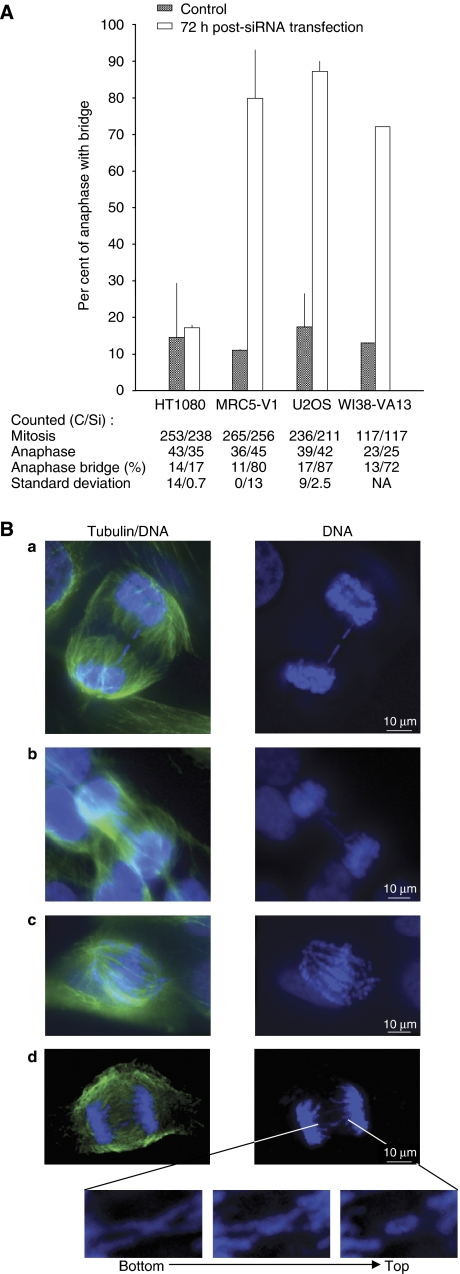
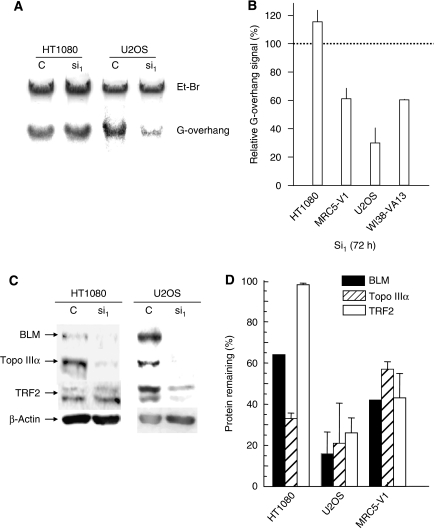
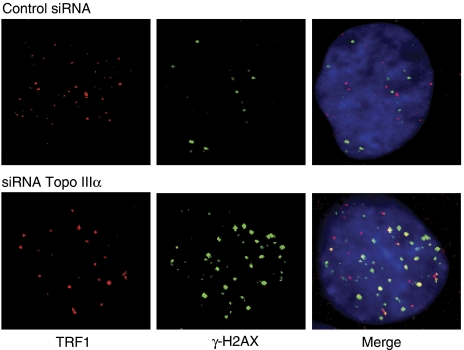
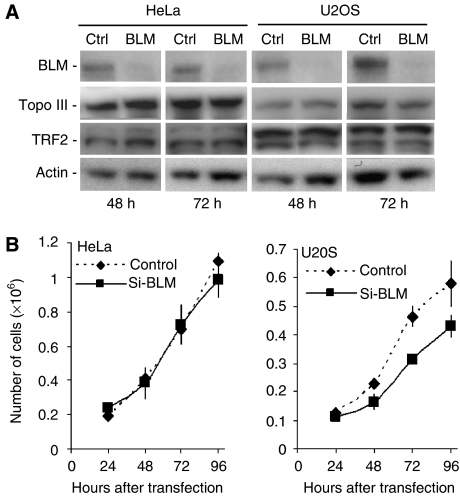
Topoisomerase (Topo) IIIalpha associates with BLM helicase, which is proposed to be important in the alternative lengthening of telomeres (ALT) pathway that allows telomere recombination in the absence of telomerase. Here, we show that human Topo IIIalpha colocalizes with telomeric proteins at ALT-associated promyelocytic bodies from ALT cells. In these cells, Topo IIIalpha immunoprecipitated with telomere binding protein (TRF) 2 and BLM and was shown to be associated with telomeric DNA by chromatin immunoprecipitation, suggesting that these proteins form a complex at telomere sequences. Topo IIIalpha depletion by small interfering RNA reduced ALT cell survival, but did not affect telomerase-positive cell lines. Moreover, repression of Topo IIIalpha expression in ALT cells reduced the levels of TRF2 and BLM proteins, provoked a strong increase in the formation of anaphase bridges, induced the degradation of the G-overhang signal, and resulted in the appearance of DNA damage at telomeres. In contrast, telomere maintenance and TRF2 levels were unaffected in telomerase-positive cells. We conclude that Topo IIIalpha is an important telomere-associated factor, essential for telomere maintenance and chromosome stability in ALT cells, and speculate on its potential mechanistic function.
Figures
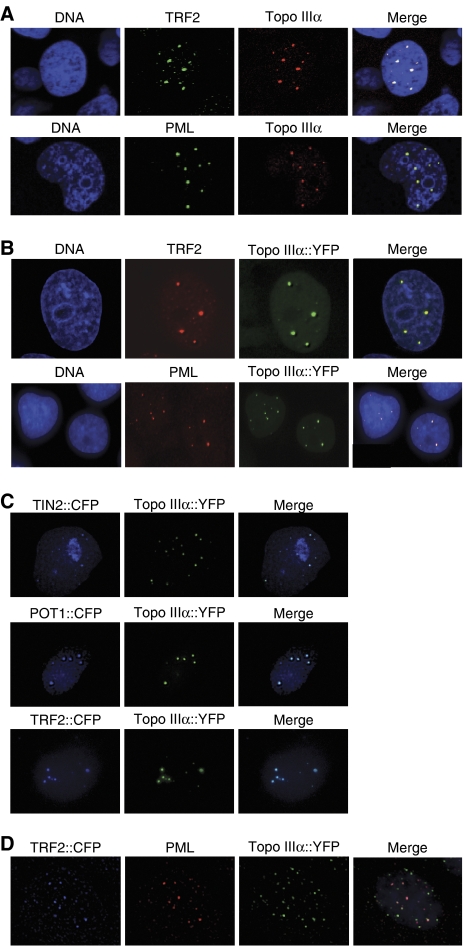
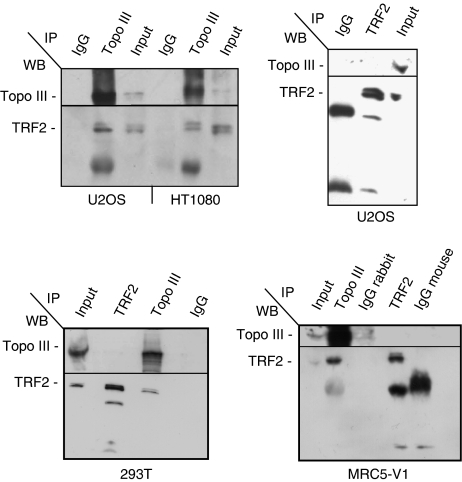
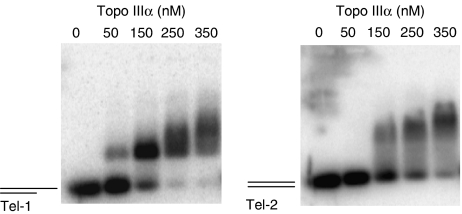
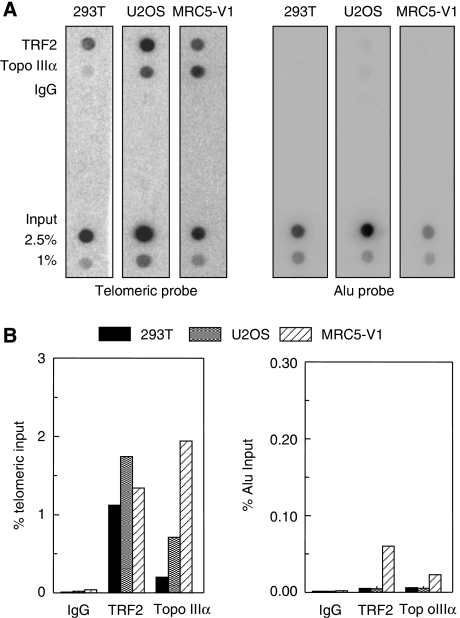
References
-
- Ababou M, Dutertre S, Lecluse Y, Onclercq R, Chatton B, Amor-Gueret M (2000) ATM-dependent phosphorylation and accumulation of endogenous BLM protein in response to ionizing radiation. Oncogene 19: 5955–5963 - PubMed
-
- Amiard S, Doudeau M, Pinte S, Poulet A, Lenain C, Faivre-Moskalenko C, Angelov D, Hug N, Vindigni A, Bouvet P, Paoletti J, Gilson E, Giraud-Panis MJ (2007) A topological mechanism for TRF2-enhanced strand invasion. Nat Struct Mol Biol 14: 147–154 - PubMed
-
- Amor-Gueret M (2006) Bloom syndrome, genomic instability and cancer: the SOS-like hypothesis. Cancer Lett 236: 1–12 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
