

Wnt5a control of cell polarity and directional movement by polarized redistribution of adhesion receptors
- PMID: 18420933
- PMCID: PMC3229220
- DOI: 10.1126/science.1151250
Wnt5a control of cell polarity and directional movement by polarized redistribution of adhesion receptors
Abstract
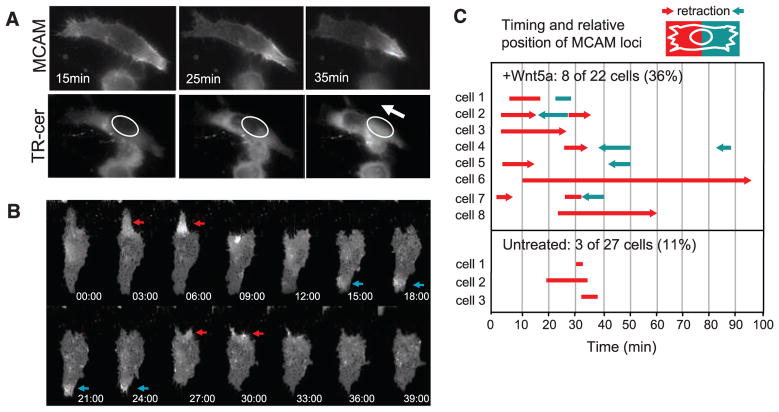
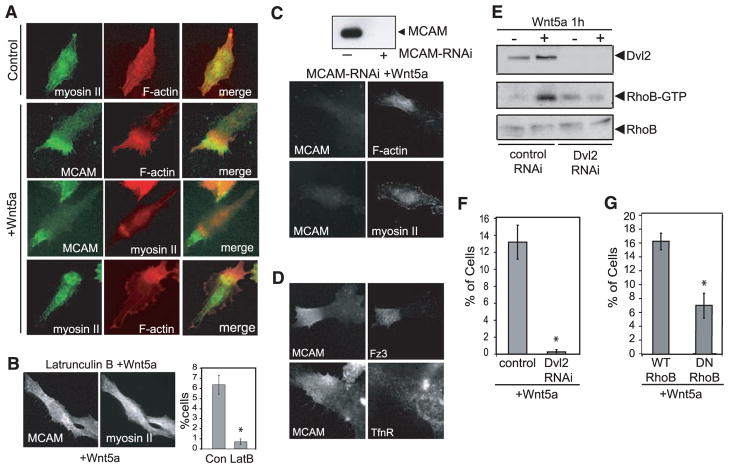
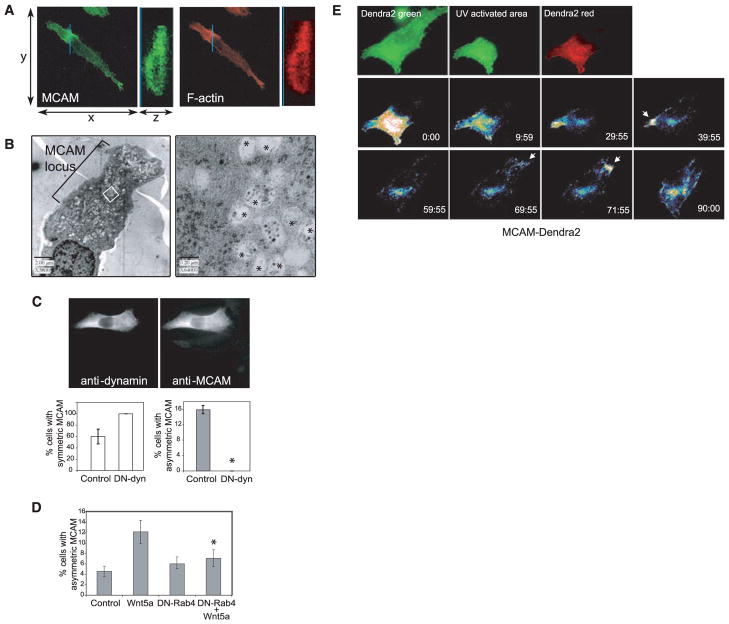
Mechanisms by which Wnt pathways integrate the organization of receptors, organelles, and cytoskeletal proteins to confer cell polarity and directional cell movement are incompletely understood. We show that acute responses to Wnt5a involve recruitment of actin, myosin IIB, Frizzled 3, and melanoma cell adhesion molecule into an intracellular structure in a melanoma cell line. In the presence of a chemokine gradient, this Wnt-mediated receptor-actin-myosin polarity (W-RAMP) structure accumulates asymmetrically at the cell periphery, where it triggers membrane contractility and nuclear movement in the direction of membrane retraction. The process requires endosome trafficking, is associated with multivesicular bodies, and is regulated by Wnt5a through the small guanosine triphosphatases Rab4 and RhoB. Thus, cell-autonomous mechanisms allow Wnt5a to control cell orientation, polarity, and directional movement in response to positional cues from chemokine gradients.
Figures

Comment in
-
Cell signaling. Wnt moves beyond the canon.Science. 2008 Apr 18;320(5874):327-8. doi: 10.1126/science.1157590. Science. 2008. PMID: 18420922 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
