

Polychaetoid controls patterning by modulating adhesion in the Drosophila pupal retina
- PMID: 18423436
- PMCID: PMC2475588
- DOI: 10.1016/j.ydbio.2008.02.022
Polychaetoid controls patterning by modulating adhesion in the Drosophila pupal retina
Abstract
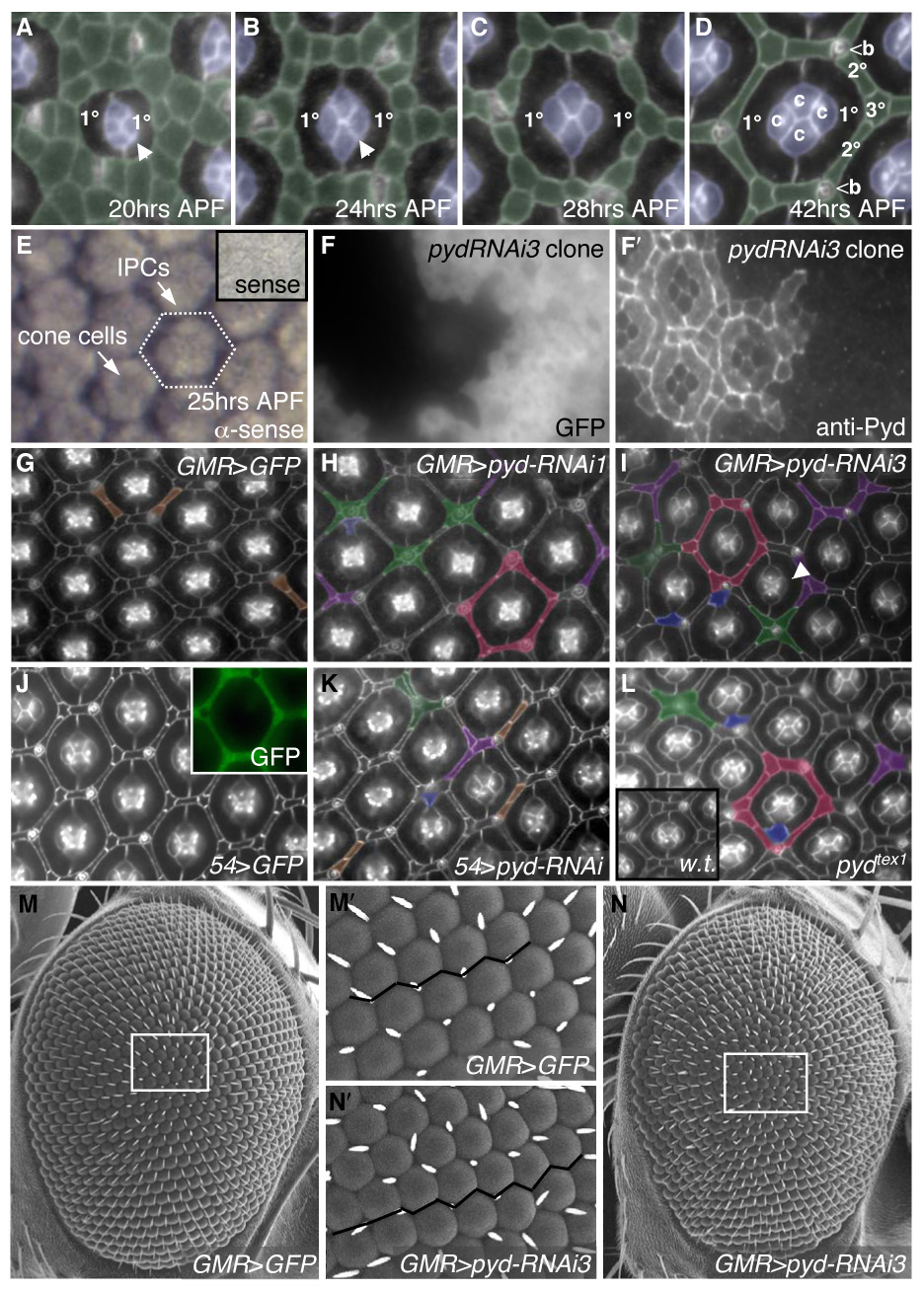
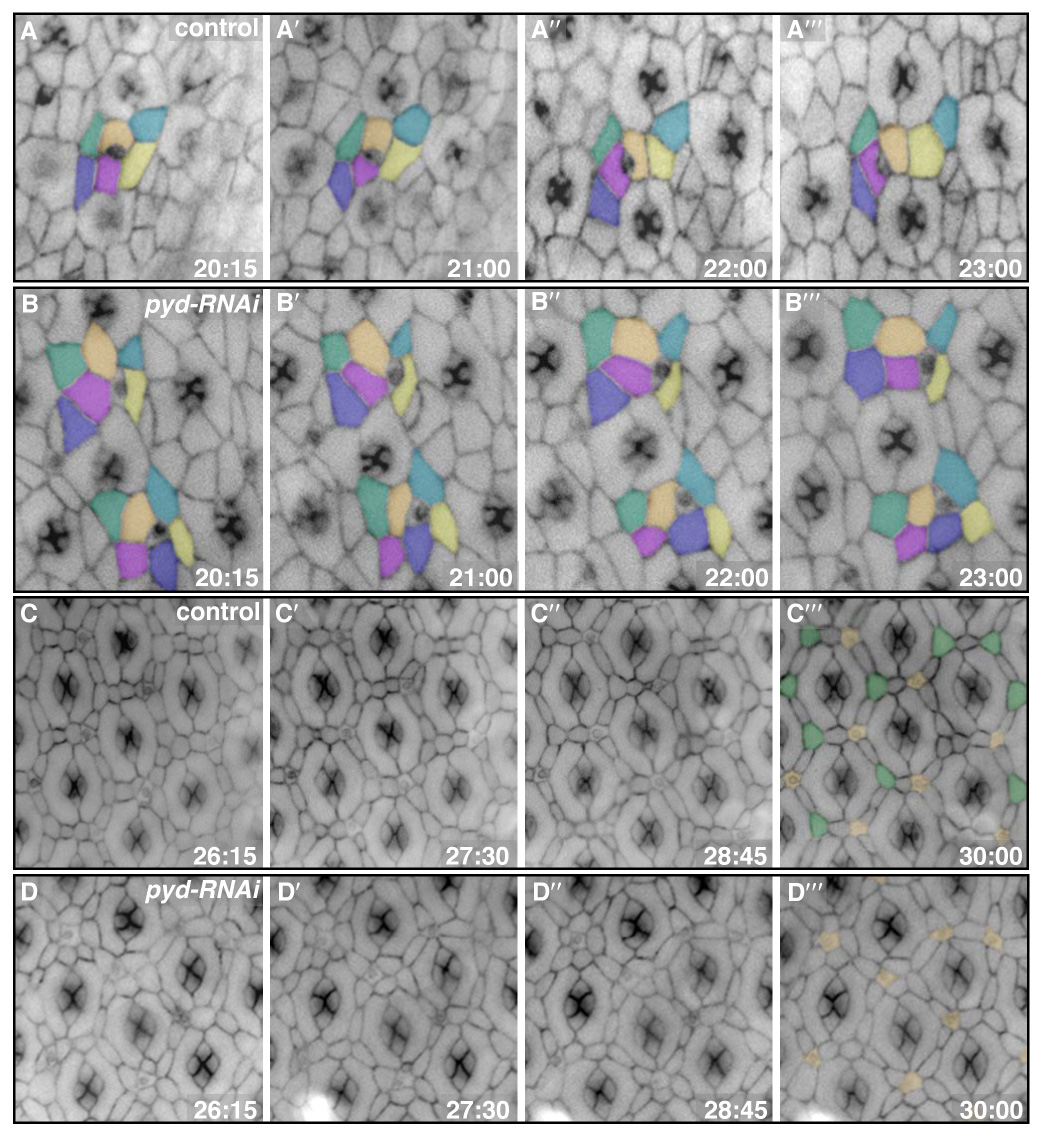
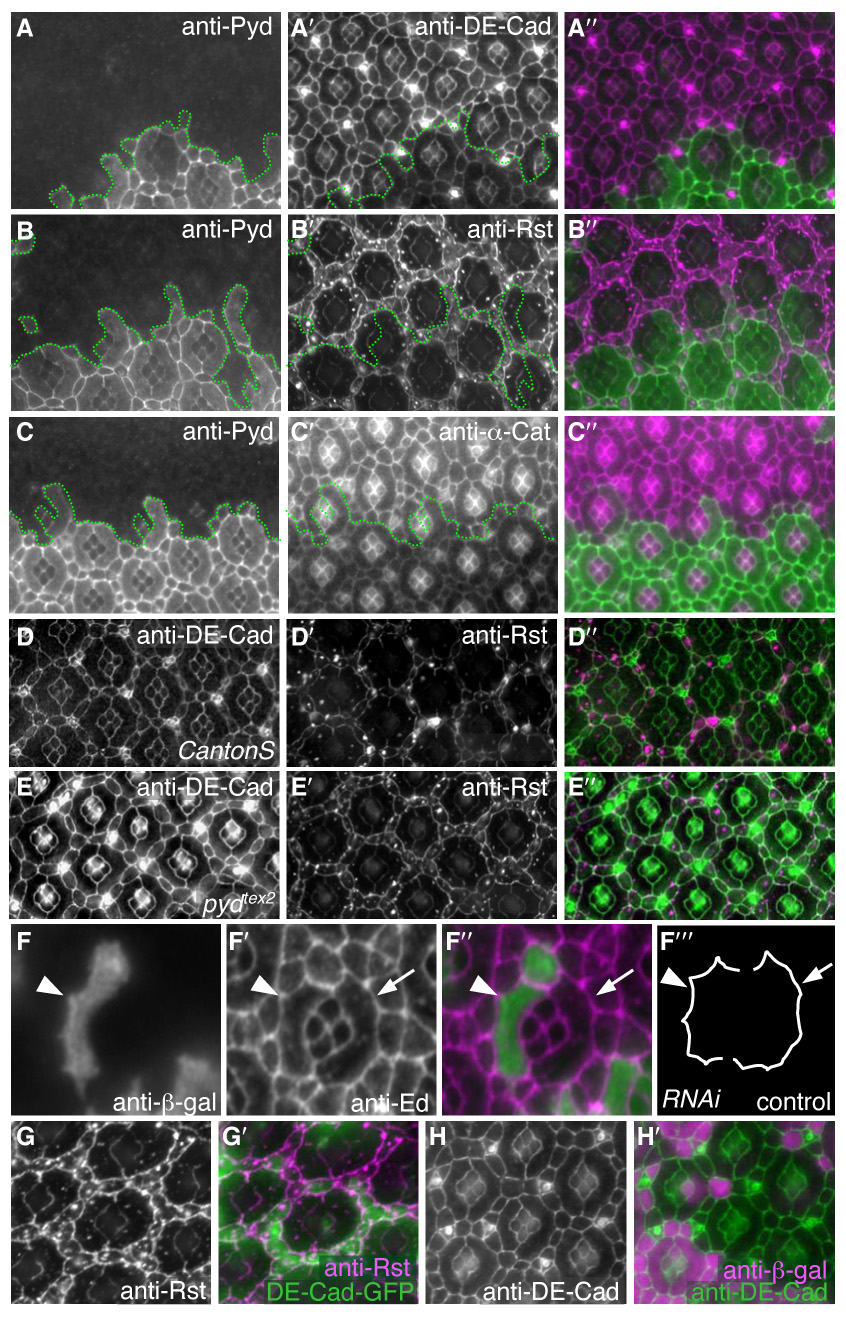
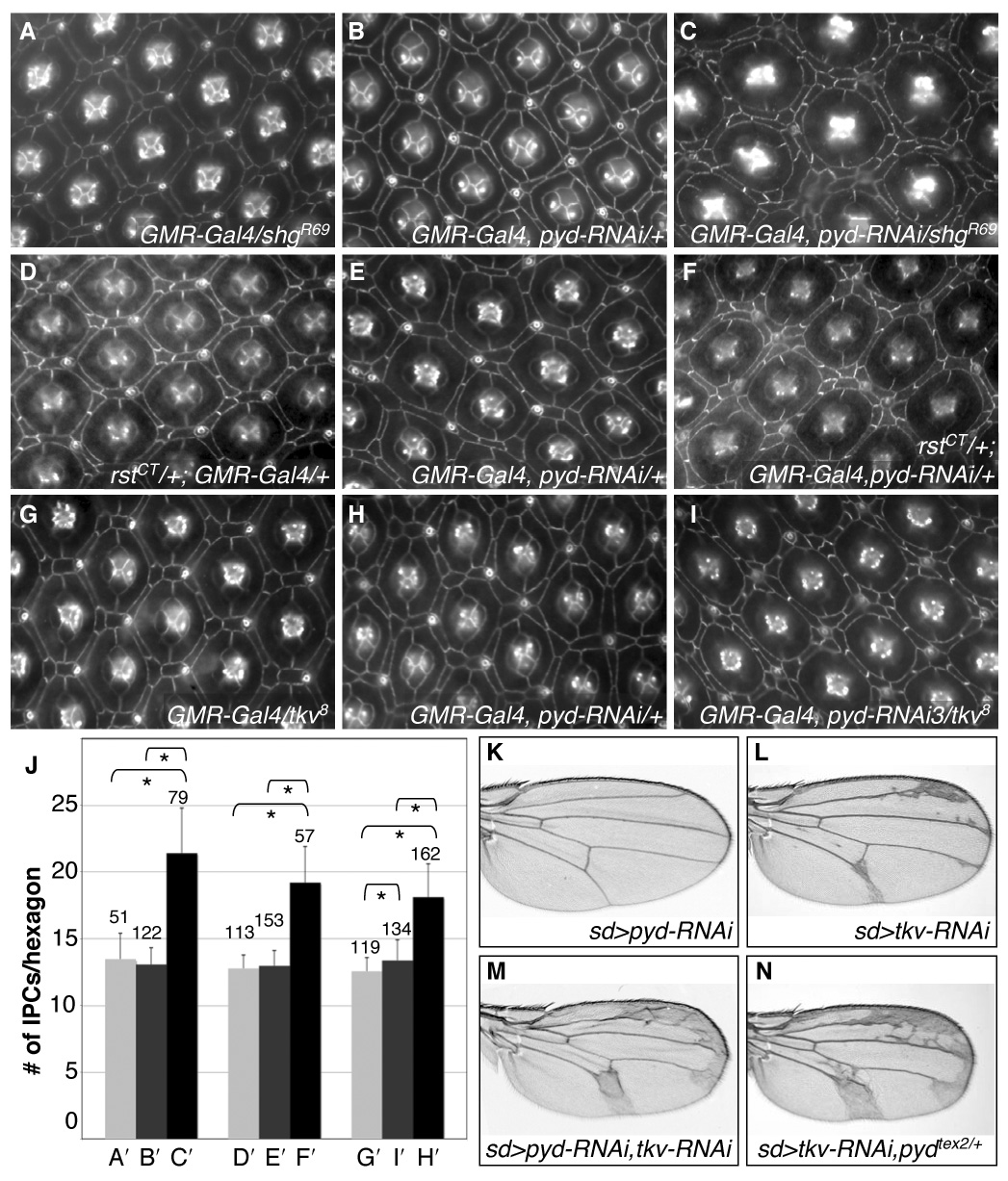
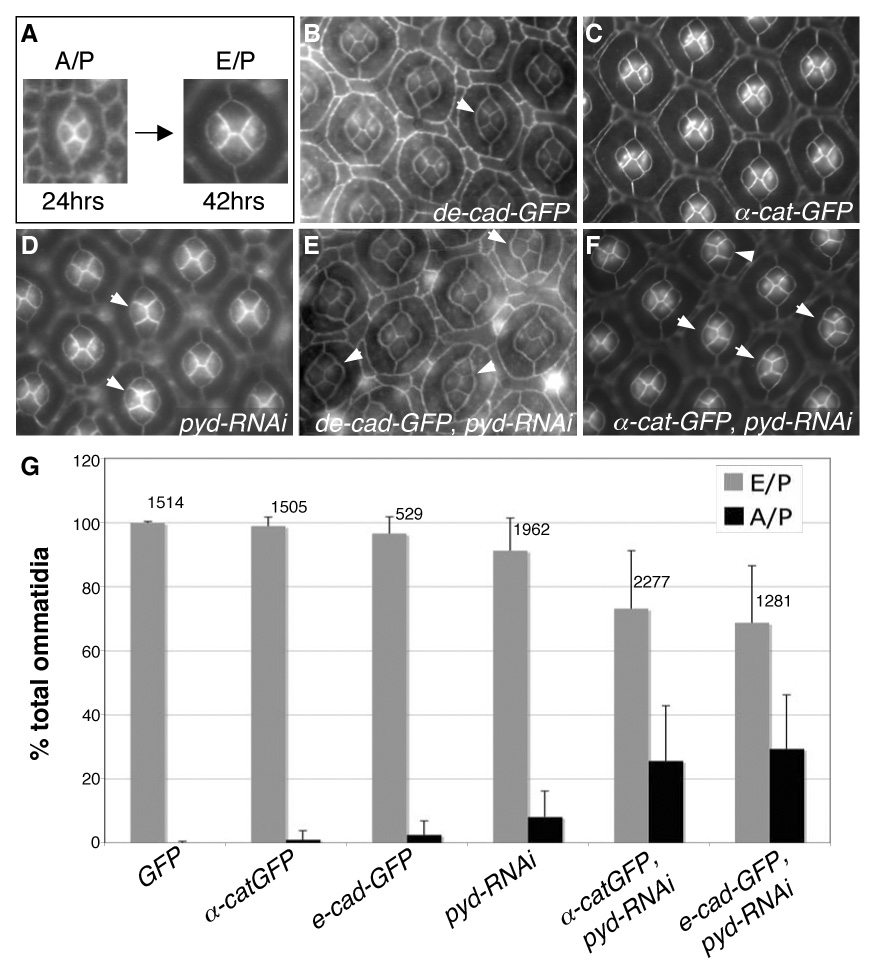
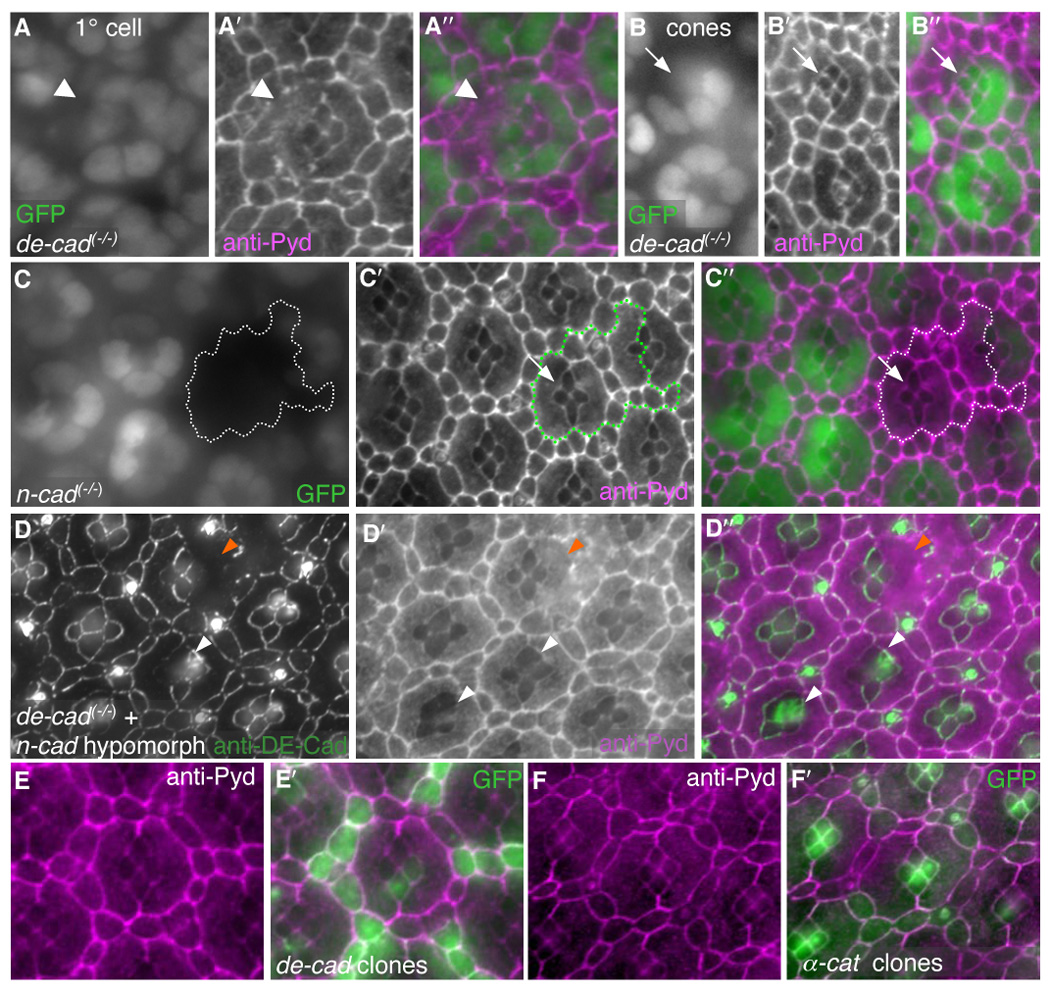
Correct cellular patterning is central to tissue morphogenesis, but the role of epithelial junctions in this process is not well-understood. The Drosophila pupal eye provides a sensitive and accessible model for testing the role of junction-associated proteins in cells that undergo dynamic and coordinated movements during development. Mutations in polychaetoid (pyd), the Drosophila homologue of Zonula Occludens-1, are characterized by two phenotypes visible in the adult fly: increased sensory bristle number and the formation of a rough eye produced by poorly arranged ommatidia. We found that Pyd was localized to the adherens junction in cells of the developing pupal retina. Reducing Pyd function in the pupal eye resulted in mis-patterning of the interommatidial cells and a failure to consistently switch cone cell contacts from an anterior-posterior to an equatorial-polar orientation. Levels of Roughest, DE-Cadherin and several other adherens junction-associated proteins were increased at the membrane when Pyd protein was reduced. Further, both over-expression and mutations in several junction-associated proteins greatly enhanced the patterning defects caused by reduction of Pyd. Our results suggest that Pyd modulates adherens junction strength and Roughest-mediated preferential cell adhesion.
Figures


References
-
- Aijaz S, Balda MS, Matter K. Tight junctions: molecular architecture and function. Int Rev Cytol. 2006;248:261–298. - PubMed
-
- Bao S, Cagan R. Preferential adhesion mediated by Hibris and Roughest regulates morphogenesis and patterning in the Drosophila eye. Dev Cell. 2005;8:925–935. - PubMed
-
- Barletta GM, Kovari IA, Verma RK, Kerjaschki D, Holzman LB. Nephrin and Neph1 co-localize at the podocyte foot process intercellular junction and form cis hetero-oligomers. J Biol Chem. 2003;278:19266–19271. - PubMed
-
- Brachmann CB, Cagan RL. Patterning the fly eye: the role of apoptosis. Trends Genet. 2003;19:91–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
