

Anaerobic and aerobic degradation of cyanophycin by the denitrifying bacterium Pseudomonas alcaligenes strain DIP1 and role of three other coisolates in a mixed bacterial consortium
- PMID: 18424548
- PMCID: PMC2423017
- DOI: 10.1128/AEM.02575-07
Anaerobic and aerobic degradation of cyanophycin by the denitrifying bacterium Pseudomonas alcaligenes strain DIP1 and role of three other coisolates in a mixed bacterial consortium
Abstract
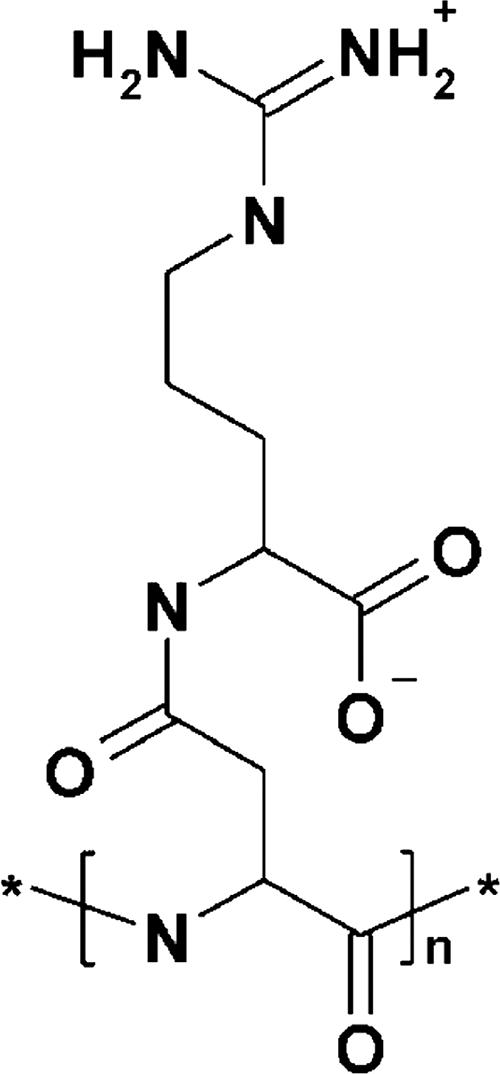
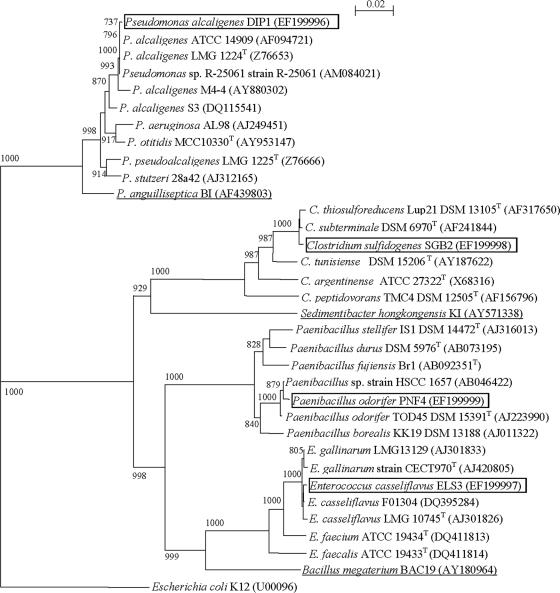
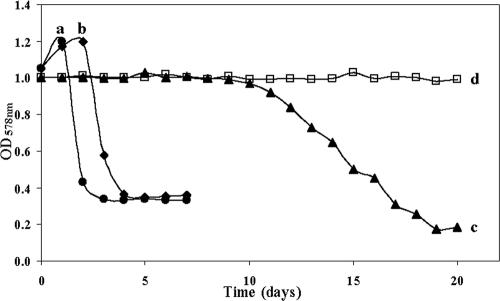
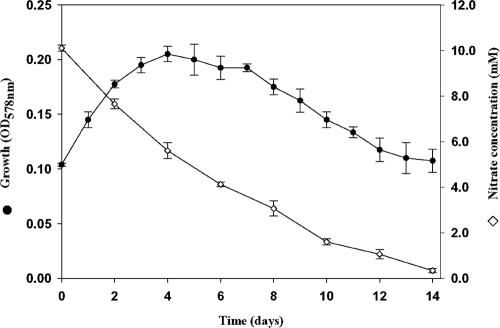
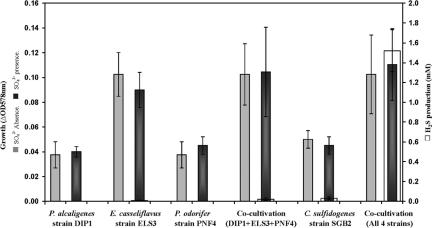
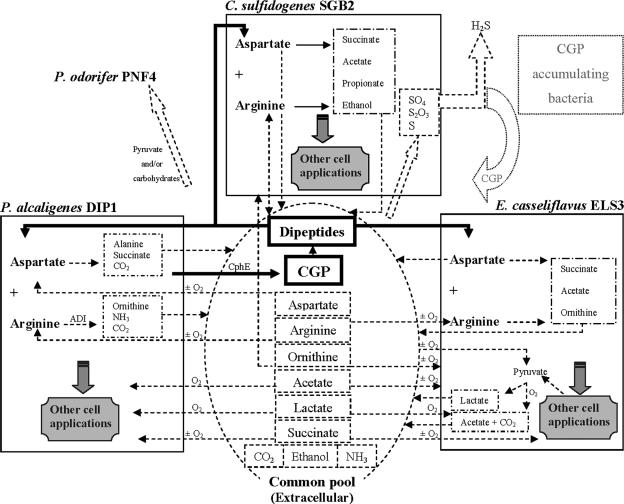
Four bacterial strains were isolated from a cyanophycin granule polypeptide (CGP)-degrading anaerobic consortium, identified by 16S rRNA gene sequencing, and assigned to species of the genera Pseudomonas, Enterococcus, Clostridium, and Paenibacillus. The consortium member responsible for CGP degradation was assigned as Pseudomonas alcaligenes strain DIP1. The growth of and CGP degradation by strain DIP1 under anaerobic conditions were enhanced but not dependent on the presence of nitrate as an electron acceptor. CGP was hydrolyzed to its constituting beta-Asp-Arg dipeptides, which were then completely utilized within 25 and 4 days under anaerobic and aerobic conditions, respectively. The end products of CGP degradation by strain DIP1 were alanine, succinate, and ornithine as determined by high-performance liquid chromatography analysis. The facultative anaerobic Enterococcus casseliflavus strain ELS3 and the strictly anaerobic Clostridium sulfidogenes strain SGB2 were coisolates and utilized the beta-linked isodipeptides from the common pool available to the mixed consortium, while the fourth isolate, Paenibacillus odorifer strain PNF4, did not play a direct role in the biodegradation of CGP. Several syntrophic interactions affecting CGP degradation, such as substrate utilization, the reduction of electron acceptors, and aeration, were elucidated. This study demonstrates the first investigation of CGP degradation under both anaerobic and aerobic conditions by one bacterial strain, with regard to the physiological role of other bacteria in a mixed consortium.
Figures
References
-
- Aboulmagd, E., F. B. Oppermann-Sanio, and A. Steinbüchel. 2000. Molecular characterization of the cyanophycin synthetase from Synechocystis sp. strain PCC6308. Arch. Microbiol. 174:297-306. - PubMed
-
- Achouak, W., P. Normand, and T. Heulin. 1999. Comparative phylogeny of rrs and nifH genes in the Bacillaceae. Int. J. Syst. Bacteriol. 49:961-967. - PubMed
-
- Appanna, V. D., R. Hamel, C. Mackenzie, P. Kumar, and S. Kalyuzhnyi. 2003. Adaptation of Pseudomonas fluorescens to Al-citrate: involvement of tricarboxylic acid and glyoxylate cycle enzymes and the influence of phosphate. Curr. Microbiol. 47:521-527. - PubMed
-
- Ariño, X., J. J. Ortega-Calvo, M. Hernandez-Marine, and C. Saiz-Jimenez. 1995. Effect of sulfur starvation on the morphology and ultrastructure of the cyanobacterium Gloeothece sp. PCC 6909. Arch. Microbiol. 163:447-453.
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
