

Arabidopsis SCARs function interchangeably to meet actin-related protein 2/3 activation thresholds during morphogenesis
- PMID: 18424615
- PMCID: PMC2390748
- DOI: 10.1105/tpc.107.055350
Arabidopsis SCARs function interchangeably to meet actin-related protein 2/3 activation thresholds during morphogenesis
Abstract
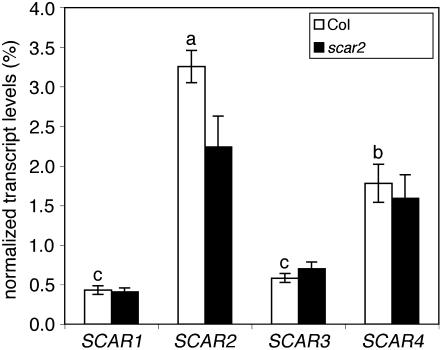
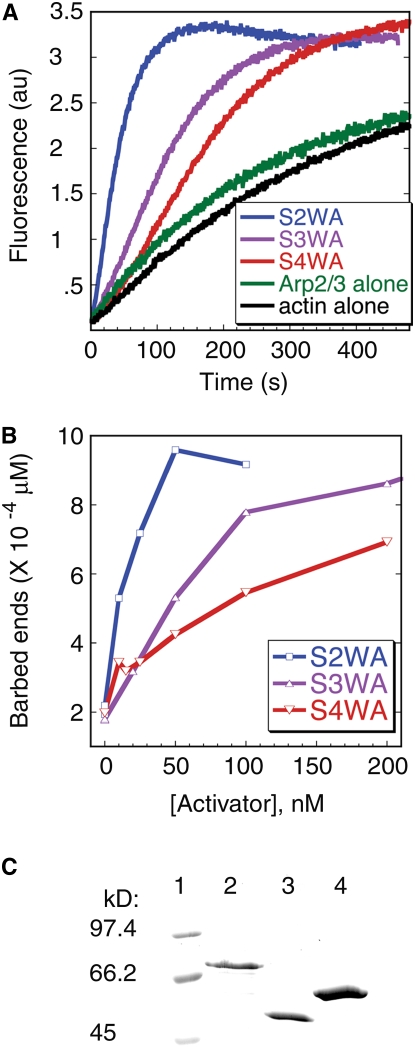
During polarized growth and tissue morphogenesis, cells must reorganize their cytoplasm and change shape in response to growth signals. Dynamic polymerization of actin filaments is one cellular component of polarized growth, and the actin-related protein 2/3 (ARP2/3) complex is an important actin filament nucleator in plants. ARP2/3 alone is inactive, and the Arabidopsis thaliana WAVE complex translates Rho-family small GTPase signals into an ARP2/3 activation response. The SCAR subunit of the WAVE complex is the primary activator of ARP2/3, and plant and vertebrate SCARs are encoded by a small gene family. However, it is unclear if SCAR isoforms function interchangeably or if they have unique properties that customize WAVE complex functions. We used the Arabidopsis distorted group mutants and an integrated analysis of SCAR gene and protein functions to address this question directly. Genetic results indicate that each of the four SCARs functions in the context of the WAVE-ARP2/3 pathway and together they define the lone mechanism for ARP2/3 activation. Genetic interactions among the scar mutants and transgene complementation studies show that the activators function interchangeably to meet the threshold for ARP2/3 activation in the cell. Interestingly, double, triple, and quadruple mutant analyses indicate that individual SCAR genes vary in their relative importance depending on the cell type, tissue, or organ that is analyzed. Differences among SCARs in mRNA levels and the biochemical efficiency of ARP2/3 activation may explain the functional contributions of individual genes.
Figures
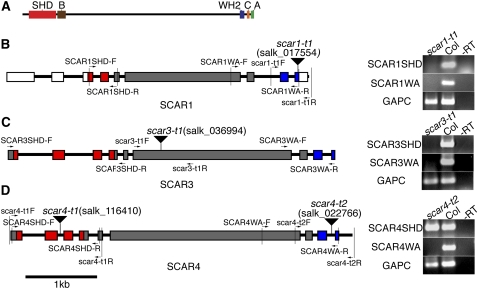
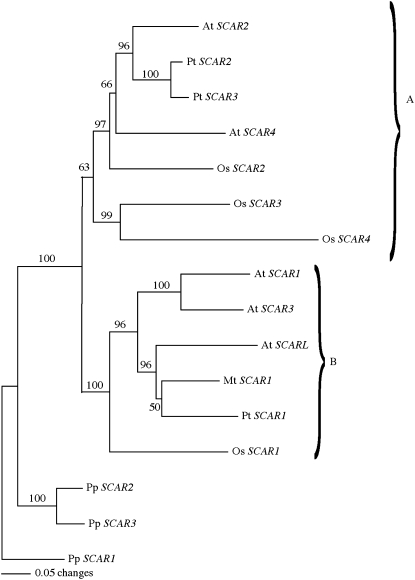
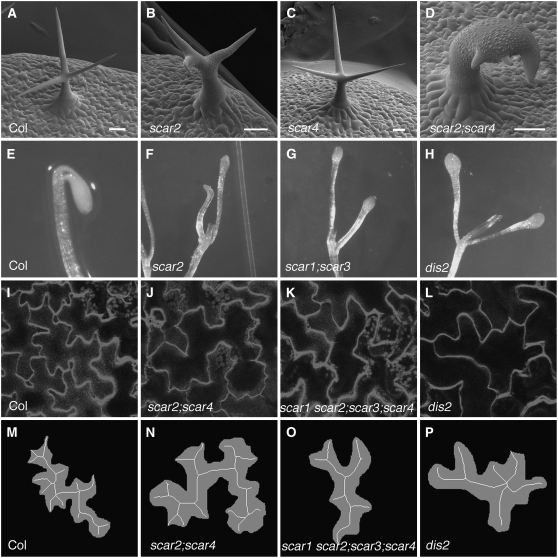
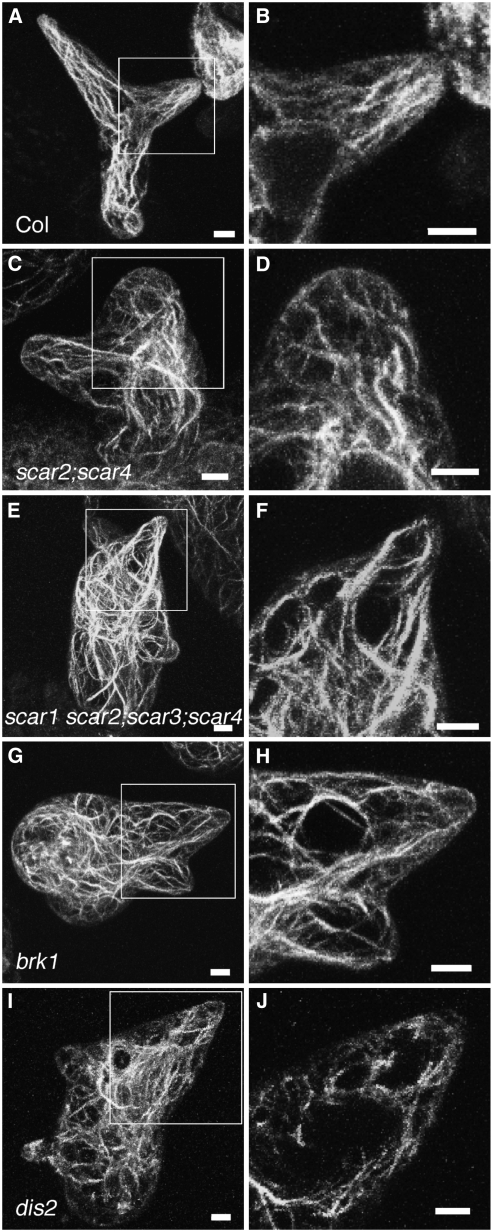

References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657. - PubMed
-
- Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol. 215 403–410. - PubMed
-
- Basu, D., El-Assal, S.E., Le, J., Mallery, E.L., and Szymanski, D.B. (2004). Interchangeable functions of Arabidopsis PIROGI and the human WAVE complex subunit SRA-1 during leaf epidermal development. Development 131 4345–4355. - PubMed
-
- Basu, D., Le, J., El-Assal, S.E., Huang, S., Zhang, C., Mallery, E.L., Koliantz, G., Staiger, C.J., and Szymanski, D.B. (2005). DISTORTED3/SCAR2 is a putative Arabidopsis WAVE complex subunit that activates the Arp2/3 complex and is required for epidermal morphogenesis. Plant Cell 17 502–524. - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
