

Prevention of allograft tolerance by bacterial infection with Listeria monocytogenes
- PMID: 18424719
- PMCID: PMC2597467
- DOI: 10.4049/jimmunol.180.9.5991
Prevention of allograft tolerance by bacterial infection with Listeria monocytogenes
Abstract
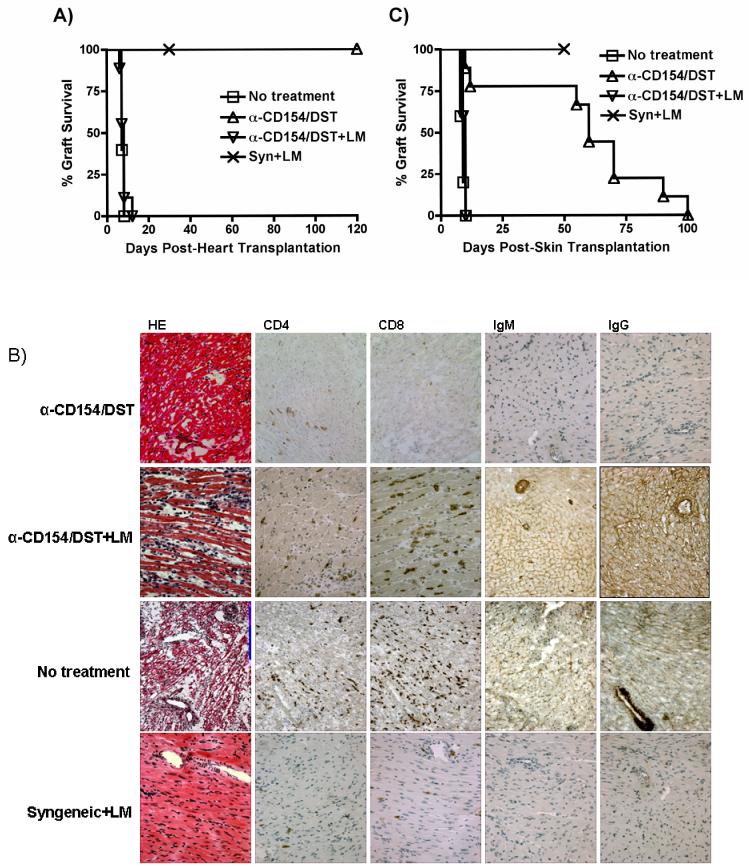
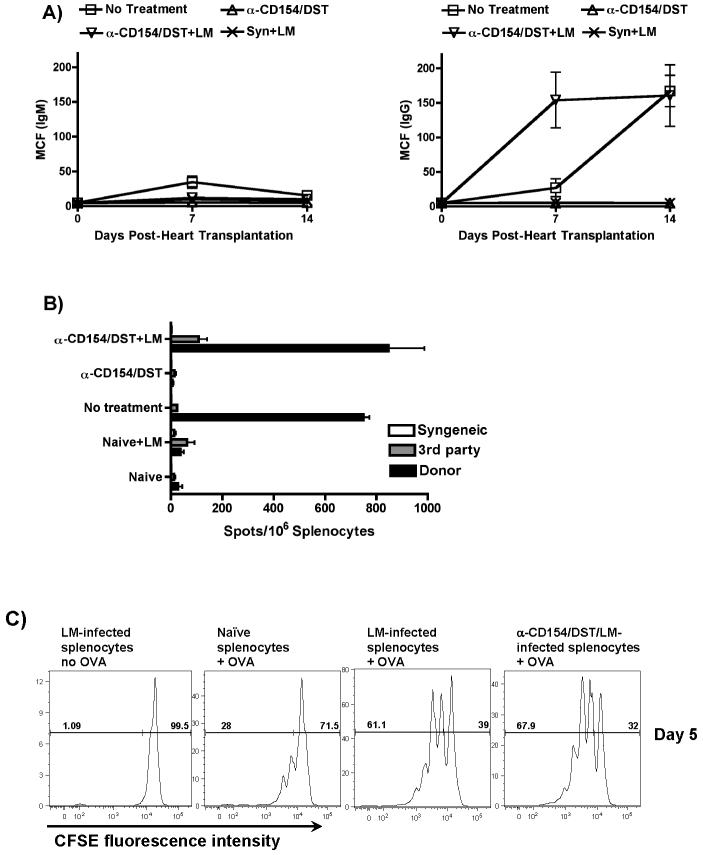
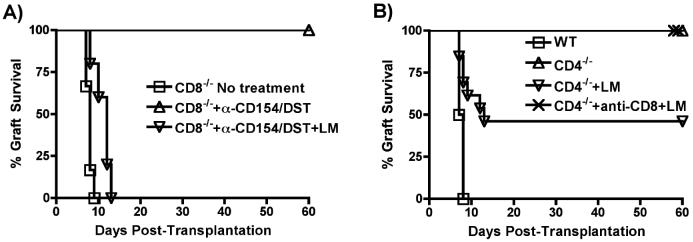
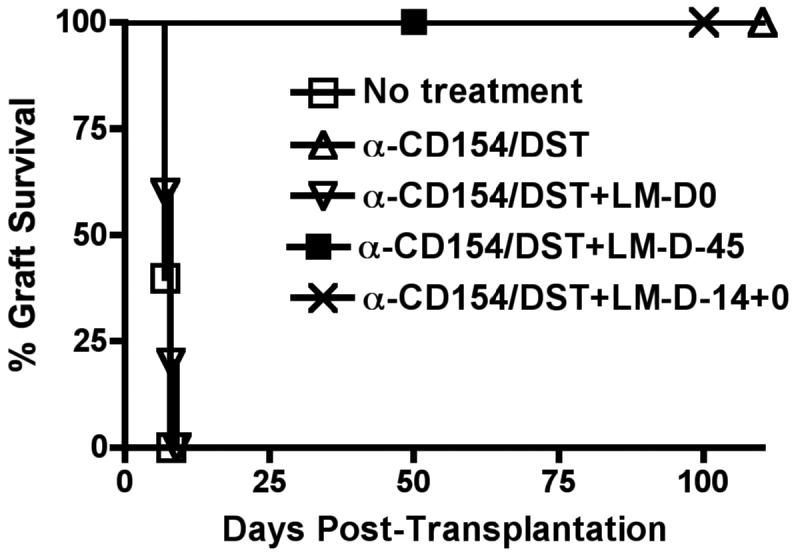
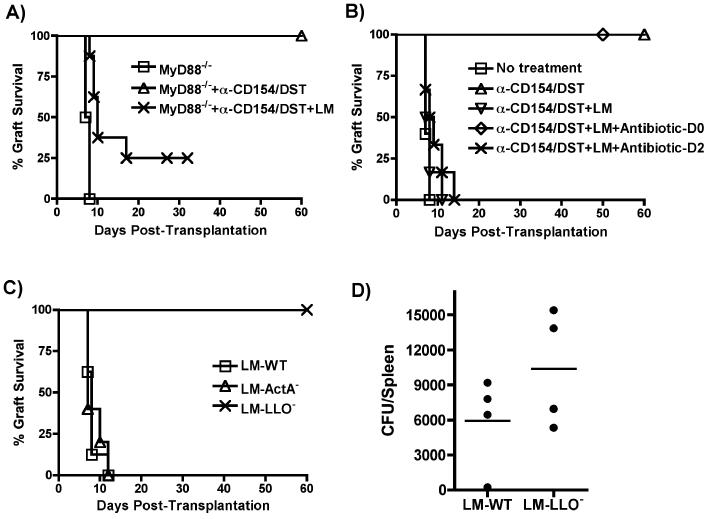
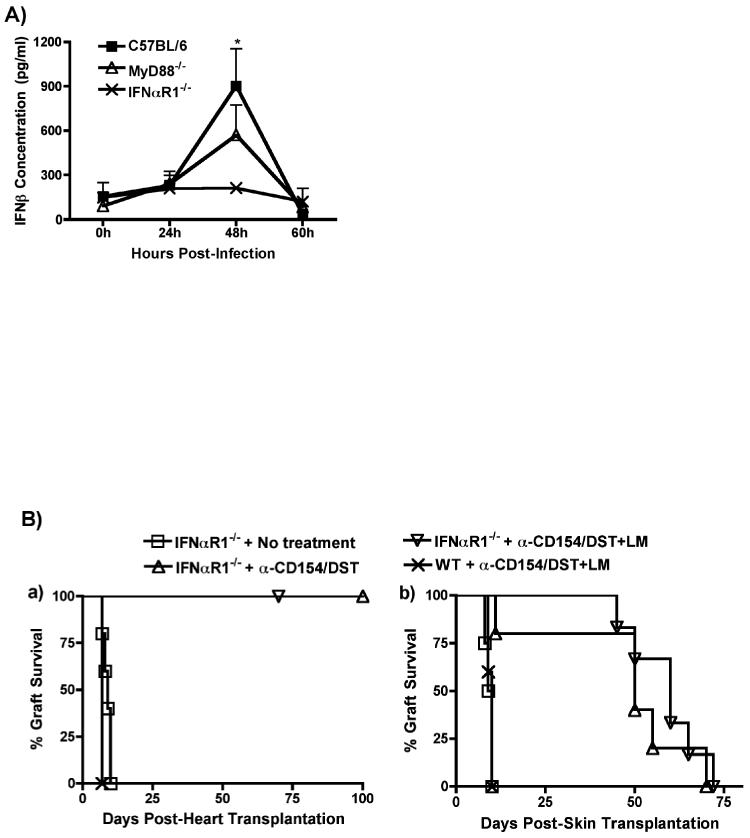
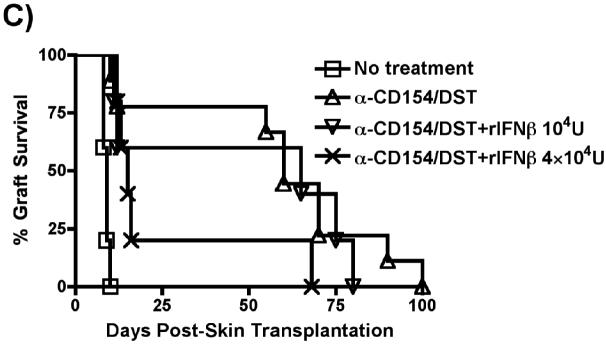
Exposure to certain viruses and parasites has been shown to prevent the induction of transplantation tolerance in mice via the generation of cross-reactive memory T cell responses or the induction of bystander activation. Bacterial infections are common in the perioperative period of solid organ allograft recipients in the clinic, and correlations between bacterial infections and acute allograft rejection have been reported. However, whether bacterial infections at the time of transplantation have any effect on the generation of transplantation tolerance remains to be established. We used the Gram-positive intracellular bacterium Listeria monocytogenes (LM) as a model pathogen because its effects on immune responses are well described. Perioperative LM infection prevented cardiac and skin allograft acceptance induced by anti-CD154 and donor-specific transfusion in mice. LM-mediated rejection was not due to the generation of cross-reactive T cells and was largely independent of signaling via MyD88, an adaptor for most TLRs, IL-1, and IL-18. Instead, transplant rejection following LM infection was dependent on the expression of the phagosome-lysing pore former listeriolysin O and on type I IFN receptor signaling. Our results indicate that bacterial exposure at the time of transplantation can antagonize tolerogenic regimens by enhancing alloantigen-specific immune responses independently of the generation of cross-reactive memory T cells.
Figures
Similar articles
-
Infection with the intracellular bacterium, Listeria monocytogenes, overrides established tolerance in a mouse cardiac allograft model.Am J Transplant. 2010 Jul;10(7):1524-33. doi: 10.1111/j.1600-6143.2010.03066.x. Am J Transplant. 2010. PMID: 20642679 Free PMC article.
-
Activation of cytosolic phospholipase A2alpha in resident peritoneal macrophages by Listeria monocytogenes involves listeriolysin O and TLR2.J Biol Chem. 2008 Feb 22;283(8):4744-55. doi: 10.1074/jbc.M709956200. Epub 2007 Dec 14. J Biol Chem. 2008. PMID: 18083708 Free PMC article.
-
Resilience of T cell-intrinsic dysfunction in transplantation tolerance.Proc Natl Acad Sci U S A. 2019 Nov 19;116(47):23682-23690. doi: 10.1073/pnas.1910298116. Epub 2019 Nov 4. Proc Natl Acad Sci U S A. 2019. PMID: 31685610 Free PMC article.
-
Mechanisms and immunological effects of apoptosis caused by Listeria monocytogenes.Adv Immunol. 2012;113:157-74. doi: 10.1016/B978-0-12-394590-7.00001-4. Adv Immunol. 2012. PMID: 22244583 Review.
-
[Immunity in listeriosis].Zh Mikrobiol Epidemiol Immunobiol. 2000 Sep-Oct;(5):98-102. Zh Mikrobiol Epidemiol Immunobiol. 2000. PMID: 11220980 Review. Russian.
Cited by
-
Alloreactive CD8 T cells rescued from apoptosis during co-stimulation blockade by Toll-like receptor stimulation remain susceptible to Fas-induced cell death.Immunology. 2013 Apr;138(4):322-32. doi: 10.1111/imm.12044. Immunology. 2013. PMID: 23190301 Free PMC article.
-
T cell responsiveness to IL-10 defines the immunomodulatory effect of costimulation blockade via anti-CD154 and impacts transplant survival.bioRxiv [Preprint]. 2024 Jun 14:2024.06.12.598652. doi: 10.1101/2024.06.12.598652. bioRxiv. 2024. PMID: 38915537 Free PMC article. Preprint.
-
CpG oligodeoxynucleotide triggers the liver inflammatory reaction and abrogates spontaneous tolerance.Liver Transpl. 2009 Aug;15(8):915-23. doi: 10.1002/lt.21771. Liver Transpl. 2009. PMID: 19642135 Free PMC article.
-
Recent progress and new perspectives in studying T cell responses to allografts.Am J Transplant. 2010 May;10(5):1117-25. doi: 10.1111/j.1600-6143.2010.03087.x. Epub 2010 Mar 26. Am J Transplant. 2010. PMID: 20353479 Free PMC article. Review.
-
Mechanisms governing bystander activation of T cells.Front Immunol. 2024 Nov 27;15:1465889. doi: 10.3389/fimmu.2024.1465889. eCollection 2024. Front Immunol. 2024. PMID: 39669576 Free PMC article. Review.
References
-
- Reinke P, Fietze E, Ode-Hakim S, Prosch S, Lippert J, Ewert R, Volk HD. Late-acute renal allograft rejection and symptomless cytomegalovirus infection. Lancet. 1994;344:1737–1738. - PubMed
-
- McLaughlin K, Wu C, Fick G, Muirhead N, Hollomby D, Jevnikar A. Cytomegalovirus seromismatching increases the risk of acute renal allograft rejection. Transplantation. 2002;74:813–816. - PubMed
-
- Sageda S, Nordal KP, Hartmann A, Sund S, Scott H, Degre M, Foss A, Leivestad T, Osnes K, Fauchald P, Rollag H. The impact of cytomegalovirus infection and disease on rejection episodes in renal allograft recipients. Am J Transplant. 2002;2:850–856. - PubMed
-
- Vilchez RA, Dauber J, McCurry K, Iacono A, Kusne S. Parainfluenza virus infection in adult lung transplant recipients: an emergent clinical syndrome with implications on allograft function. Am J Transplant. 2003;3:116–120. - PubMed
-
- Pantenburg B, Heinzel F, Das L, Heeger PS, Valujskikh A. T cells primed by Leishmania major infection cross-react with alloantigens and alter the course of allograft rejection. J Immunol. 2002;169:3686–3693. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
