

The inositol phosphatase SHIP controls Salmonella enterica serovar Typhimurium infection in vivo
- PMID: 18426884
- PMCID: PMC2446709
- DOI: 10.1128/IAI.01596-07
The inositol phosphatase SHIP controls Salmonella enterica serovar Typhimurium infection in vivo
Abstract
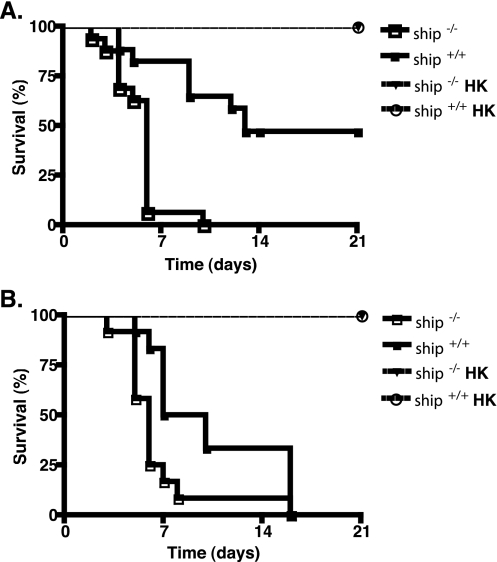
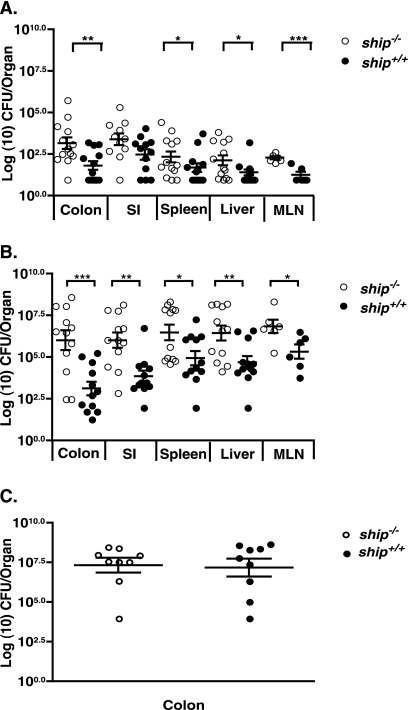
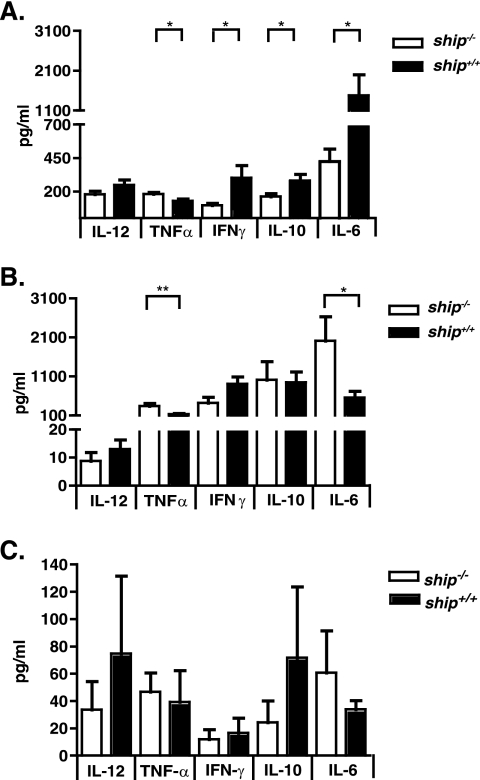
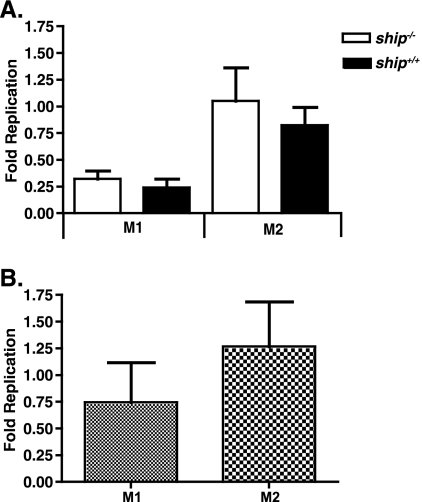
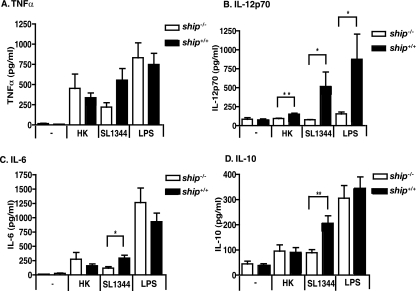
The SH2 domain-containing inositol 5'-phosphatase, SHIP, negatively regulates various hematopoietic cell functions and is critical for maintaining immune homeostasis. However, whether SHIP plays a role in controlling bacterial infections in vivo remains unknown. Salmonella enterica causes human salmonellosis, a disease that ranges in severity from mild gastroenteritis to severe systemic illness, resulting in significant morbidity and mortality worldwide. The susceptibility of ship(+/+) and ship(-/-) mice and bone marrow-derived macrophages to S. enterica serovar Typhimurium infection was compared. ship(-/-) mice displayed an increased susceptibility to both oral and intraperitoneal serovar Typhimurium infection and had significantly higher bacterial loads in intestinal and systemic sites than ship(+/+) mice, indicating a role for SHIP in the gut-associated and systemic pathogenesis of serovar Typhimurium in vivo. Cytokine analysis of serum from orally infected mice showed that ship(-/-) mice produce lower levels of Th1 cytokines than do ship(+/+) animals at 2 days postinfection, and in vitro analysis of supernatants taken from infected bone marrow-derived macrophages derived to mimic the in vivo ship(-/-) alternatively activated (M2) macrophage phenotype correlated with these data. M2 macrophages were the predominant population in vivo in both oral and intraperitoneal infections, since tissue macrophages within the small intestine and peritoneal macrophages from ship(-/-) mice showed elevated levels of the M2 macrophage markers Ym1 and Arginase 1 compared to ship(+/+) cells. Based on these data, we propose that M2 macrophage skewing in ship(-/-) mice contributes to ineffective clearance of Salmonella in vivo.
Figures


References
-
- Brauweiler, A. M., I. Tamir, and J. C. Cambier. 2000. Bilevel control of B-cell activation by the inositol 5-phosphatase SHIP. Immunol. Rev. 17669-74. - PubMed
-
- CDC. 2005. Salmonella surveillance: annual summary, 2004. U.S. Department of Health and Human Services, CDC, Atlanta, GA.
-
- Damen, J. E., L. Liu, P. Rosten, R. K. Humphries, A. B. Jefferson, P. W. Majerus, and G. Krystal. 1996. The 145-kDa protein induced to associate with Shc by multiple cytokines is an inositol tetraphosphate and phosphatidylinositol 3,4,5-triphosphate 5-phosphatase. Proc. Natl. Acad. Sci. USA 931689-1693. - PMC - PubMed
-
- Eckmann, L., and M. F. Kagnoff. 2001. Cytokines in host defense against Salmonella. Microbes Infect./Inst. Pasteur 31191-1200. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
