

Review
doi: 10.1021/cr0782067.
Epub 2008 Apr 23.
Enzymatic pathways that regulate endocannabinoid signaling in the nervous system
Affiliations
- PMID: 18429637
- PMCID: PMC3150828
- DOI: 10.1021/cr0782067
Item in Clipboard
Review
Enzymatic pathways that regulate endocannabinoid signaling in the nervous system
Chem Rev.
2008 May.
No abstract available
Figures
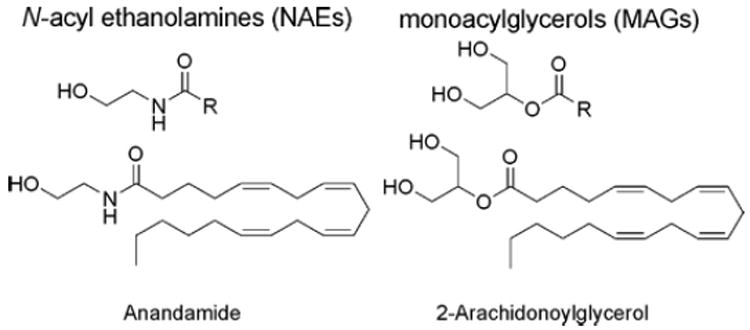
Two principle endocannabinoids, N-arachidonoyl ethanolamine (anandamide) and 2-arachidonoylglycerol (2-AG), which are members of theN-acyl ethanolamine (NAE) and monoacylglycerol (MAG) classes of lipids, respectively.
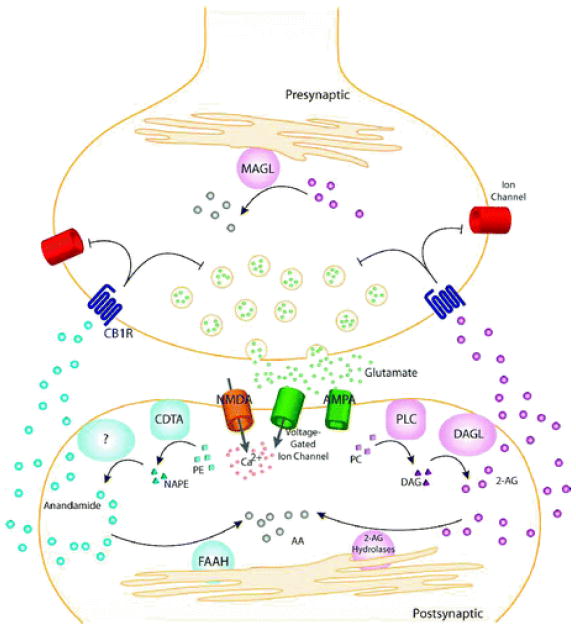
General model for endocannabinoid-based retrograde signaling. Upon release of neurotransmitter (e.g., glutamate), postsynaptic receptors (e.g., α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), N-methyl- d -aspartic acid (NMDA)) and voltage-gated ion channels are activated, allowing influx of Ca 2+ and on-demand endocannabinoid biosynthesis. Anandamide is synthesized from phospholipid precursors by a calcium-dependent transacylase (CDTA) and one or more other still uncharacterized enzymes. 2-Arachidonoylglycerol (2-AG) is synthesized from phospholipid precursors by phospholipase C (PLC) and diacylglycerol lipase (DAGL). Endocannabinoids then migrate from postsynaptic neurons to CB1 receptors (CB1R) located on presynaptic neurons. Once activated, CB1Rs couple through the G i/G o class of G-proteins to regulate ion channels and inhibit neurotransmitter release. The retrograde signaling of endocannabinoids is then terminated by degradative enzymes. Anandamide is hydrolyzed to arachidonic acid (AA) primarily by fatty acid amide hydrolase (FAAH), located in the postsynaptic neuron. 2-AG is hydrolyzed to AA primarily by monoacylglycerol lipase (MAGL) in the presynaptic neuron, though other 2-AG hydrolases may also participate in this process.
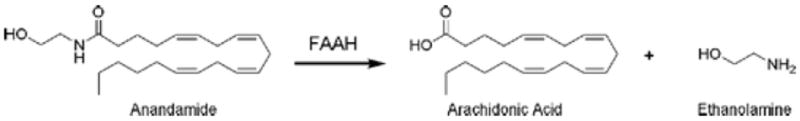
Enzymatic hydrolysis of anandamide to arachidonic acid and ethanolamine catalyzed by FAAH.
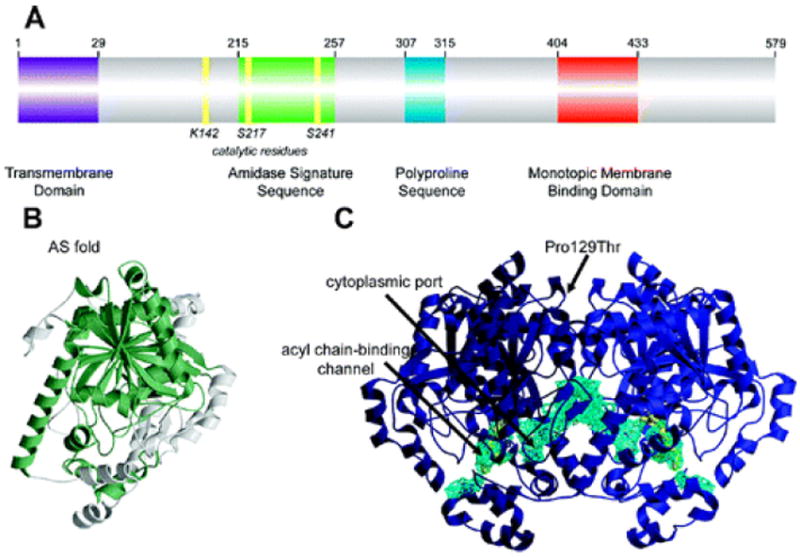
Structural features of FAAH: (A) Primary sequence analysis reveals a predicted NH 2-terminal transmembrane domain (purple), an amidase signature sequence rich in glycine and serine residues (green), a polyproline sequence predicted to interact with Homer and SH3 domain-containing proteins (blue), and a monotopic membrane binding domain that enables FAAH to bind the membrane even in the absence of the transmembrane domain (red). (B) An overlay of the known structures of amidase signature (AS) enzymes reveals a common “AS fold”, shown in green for the FAAH monomer. (C) Two channels in the FAAH X-ray crystal structure suggest possible routes for substrate binding (acyl chain-binding channel) and product release (cytoplasmic port). Structural studies also revealed that a common human single nucleotide polymorphism, which results in mutation of Pro129 to threonine, is located on the putative cytoplasmic face of FAAH.
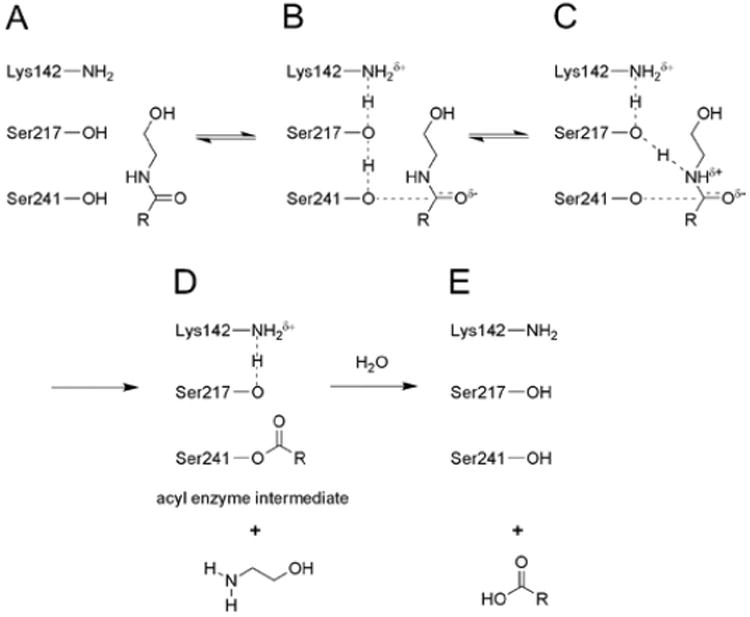
Proposed mechanism for the hydrolysis of amide and ester substrates catalyzed by FAAH (shown for amides): (A) Lys142, initially in a deprotonated state, (B) abstracts a proton from Ser217, which in turn abstracts a proton from the Ser241 nucleophile. (C) Attack of the nucleophile on the substrate carbonyl is proposed to occur in a coupled manner with proton donation from Ser217 to the nitrogen atom of the amide substrate. This latter step requires the concurrent donation of a proton from Lys142 to Ser217, resulting in (D) the formation of an acyl–enzyme intermediate where both Lys142 and Ser217 have returned to their initial protonation states. (E) Deacylation results in release of the free fatty acid product.
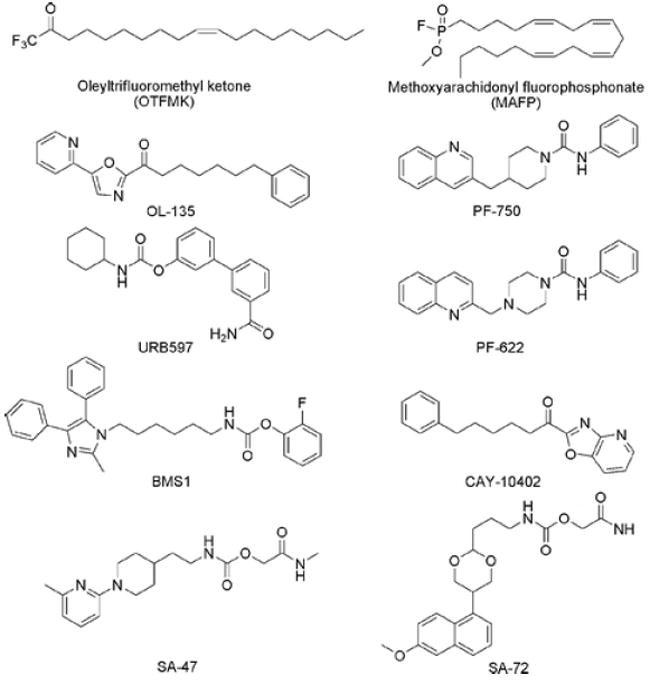
Representative FAAH inhibitors.
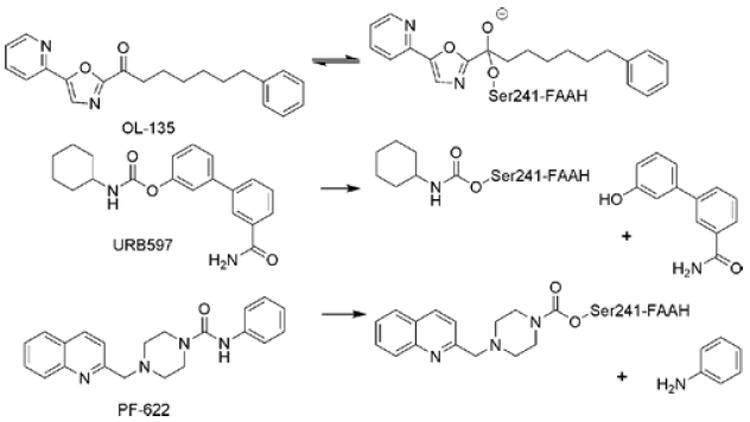
Different modes of action for classes of FAAH inhibitors. OL-135, an α-ketoheterocycle, is a covalent, reversible inhibitor of FAAH. URB597, a carbamate, and PF-622, a piperazine urea, irreversibly inhibit FAAH by carbamylation of the Ser241 nucleophile.
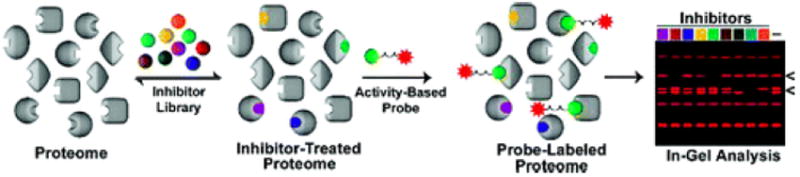
Competitive activity-based protein profiling (ABPP). To determine the selectivity of an inhibitor library against serine hydrolases, a proteome is reacted with inhibitor and subsequently labeled with a rhodamine-tagged fluorophosphonate. Reacted proteomes are then analyzed by 1-D SDS-PAGE. A decrease in fluorescent intensity of the probe in the presence of inhibitor indicates a target.
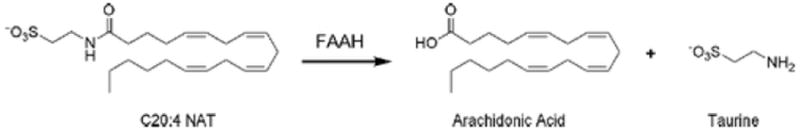
Enzymatic hydrolysis of C20:4 N-acyl taurine (NAT) to arachidonic acid and taurine catalyzed by FAAH.
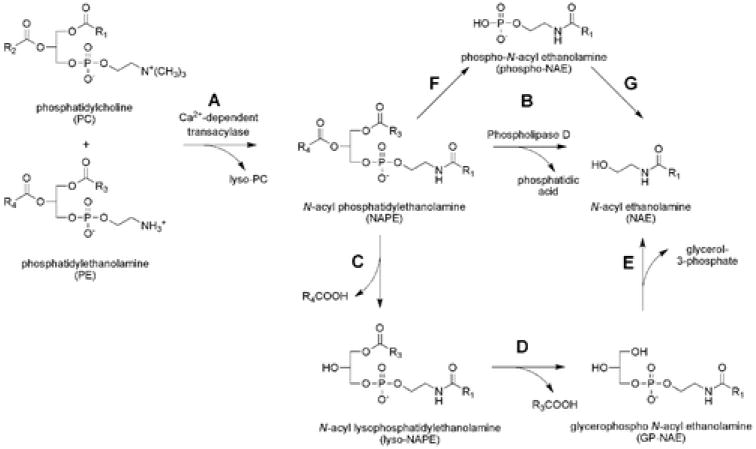
Postulated routes for the biosynthesis of anandamide and other NAEs: (A) A calcium-dependent transacylase (CDTA) converts phospholipid precursors to N-acyl phosphatidylethanolamine (NAPE). (B) Phospholipase D (PLD) then hydrolyzes NAPEs to produce NAEs. (C–E) An alternative PLD-independent pathway in which the sn-1 and sn-2 O-acyl chains of NAPEs are hydrolyzed to generate the intermediates lyso-NAPE (C) and glycerophospho (GP)-NAE (D), respectively. Subsequent cleavage of the phosphodiester bonds of lyso-NAPE and GP-NAE would then yield NAEs (E). (F, G) A third pathway for the conversion of NAPEs to NAEs involves a phospholipase C-dependent conversion of NAPEs to phospho-NAEs (F) followed by the hydrolysis of phospho-NAEs to NAEs by phosphatase-mediated hydrolysis (G).
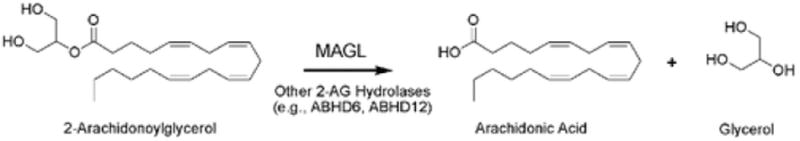
Enzymatic hydrolysis of 2-AG to arachidonic acid and glycerol catalyzed by MAGL and other hydrolases (e.g., ABHD6, ABHD12).
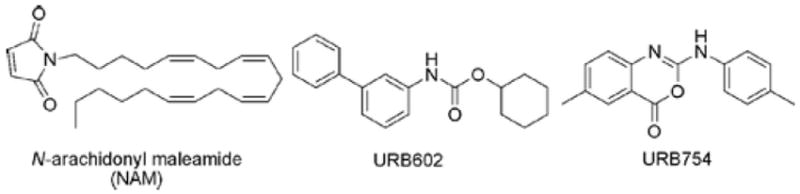
Reported MAGL inhibitors. Note that URB754 has since been shown not to inhibit MAGL.
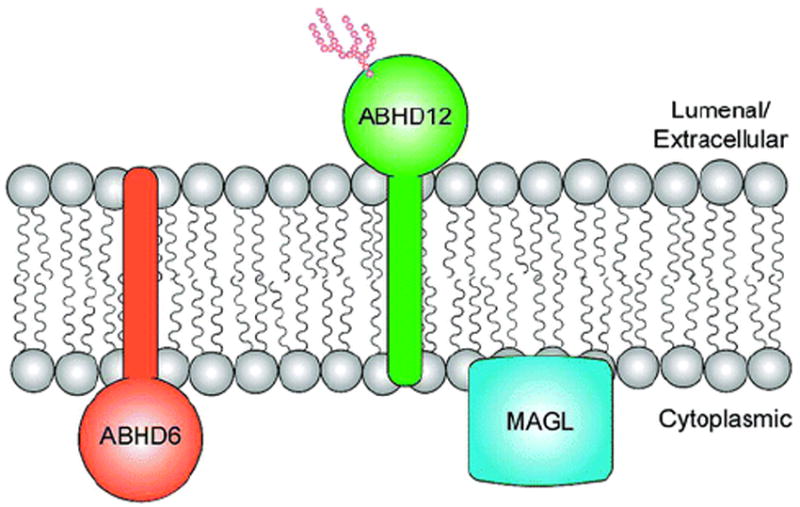
Three prominent 2-AG hydrolases in the nervous system, MAGL, ABHD6, and ABHD12, have different subcellular distributions, suggesting that they could degrade distinct pools of 2-AG.
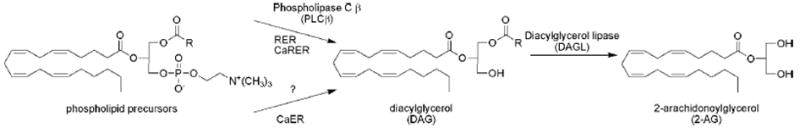
Enzymatic biosynthesis of 2-AG. Receptor-regulated endocannabinoid release (RER) and calcium-assisted RER (CaRER) employ a phospholipase C β (PLCβ)-dependent pathway to convert phospholipid precursors to diacylglycerol (DAG). The source of DAG for calcium-dependent endocannabinoid release (CaER) is currently unknown. Both pathways intersect with the formation of DAG, which is then converted to 2-AG precursors by diacylglycerol lipases (DAGL-α and -β).
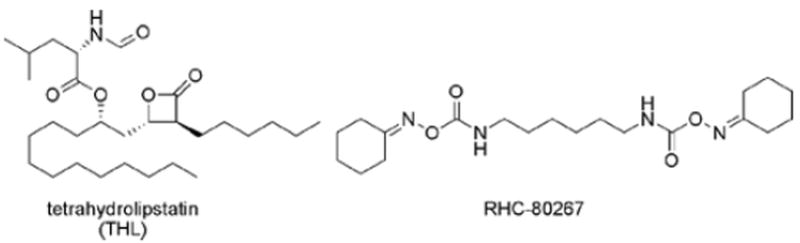
Representative DAGLα and -β inhibitors.
References
-
- Siegel GJ, Agranoff BW, Albers RW, Molinoff PB. Basic Neurochemistry. Raven Press; New York: 1994.
-
- Mechoulam R. The Pharmacohistory of Cannabis sativa. CRC Press; Boca Raton, FL: 1986.
-
- Mackie K. Annu Rev Pharmacol Toxicol. 2006;46:101. - PubMed
-
- Ledent C, Valverde O, Cossu G, Petitet F, Aubert JF, B F, Bohme GA, Imperato A, Pedrazzini T, Roques BP, Vassart G, Fratta W, Parmentier M. Science. 1999;283:401. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
