

The transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements
- PMID: 18433450
- PMCID: PMC2440561
- DOI: 10.1111/j.1365-2958.2008.06249.x
The transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements
Abstract
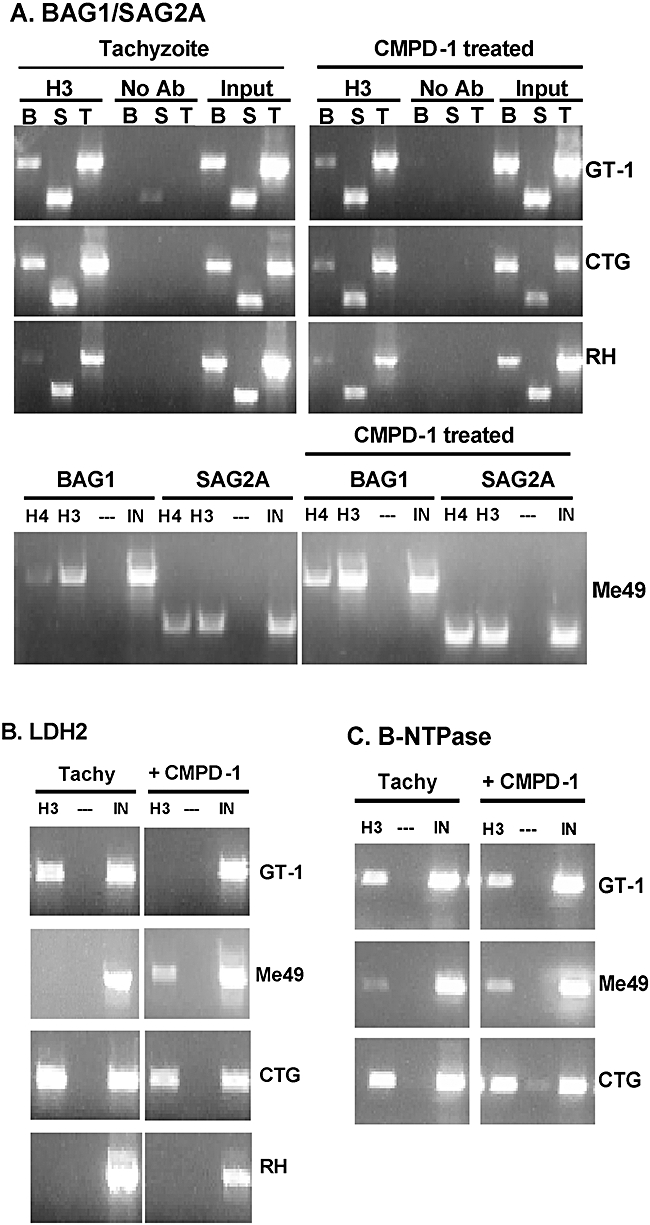
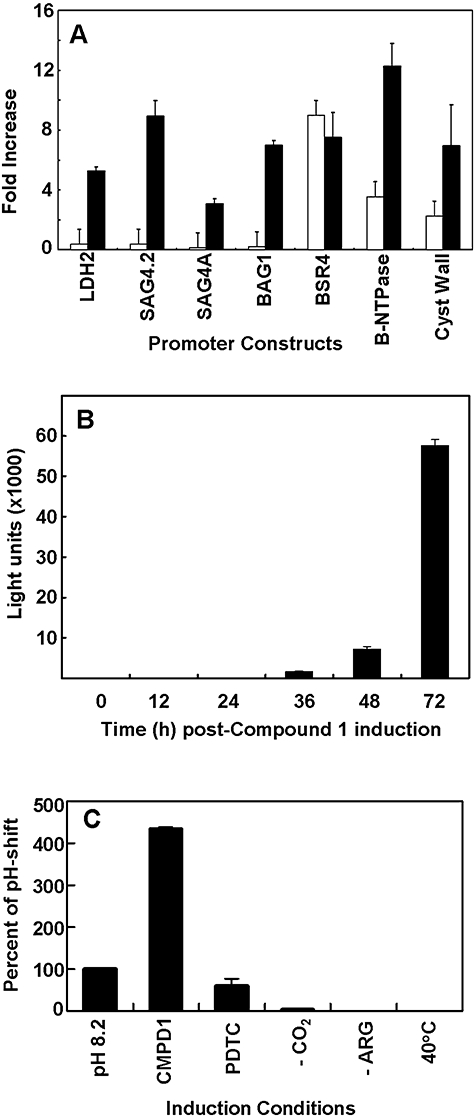
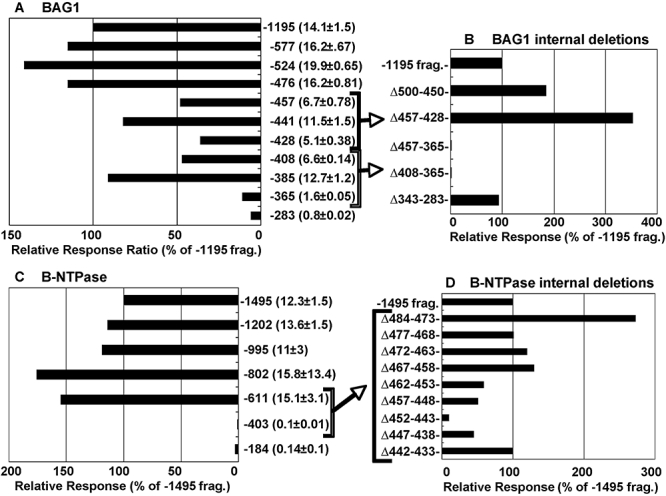
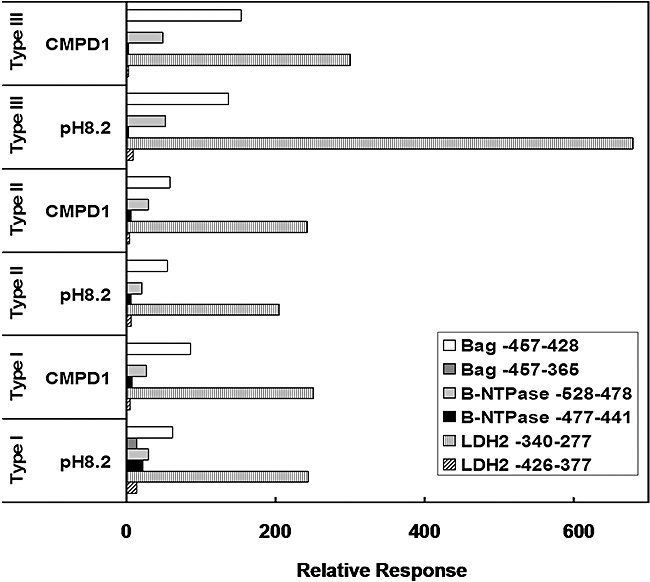
Experimental evidence suggests that apicomplexan parasites possess bipartite promoters with basal and regulated cis-elements similar to other eukaryotes. Using a dual luciferase model adapted for recombinational cloning and use in Toxoplasma gondii, we show that genomic regions flanking 16 parasite genes, which encompass examples of constitutive and tachyzoite- and bradyzoite-specific genes, are able to reproduce the appropriate developmental stage expression in a transient luciferase assay. Mapping of cis-acting elements in several bradyzoite promoters led to the identification of short sequence spans that are involved in control of bradyzoite gene expression in multiple strains and under different bradyzoite induction conditions. Promoters that regulate the heat shock protein BAG1 and a novel bradyzoite-specific NTPase during bradyzoite development were fine mapped to a 6-8 bp resolution and these minimal cis-elements were capable of converting a constitutive promoter to one that is induced by bradyzoite conditions. Gel-shift experiments show that mapped cis-elements are bound by parasite protein factors with the appropriate functional sequence specificity. These studies are the first to identify the minimal sequence elements that are required and sufficient for bradyzoite gene expression and to show that bradyzoite promoters are maintained in a 'poised' chromatin state throughout the intermediate host life cycle in low passage strains. Together, these data demonstrate that conventional eukaryotic promoter mechanisms work with epigenetic processes to regulate developmental gene expression during tissue cyst formation.
Figures




References
-
- Ajzenberg D, Banuls AL, Su C, Dumetre A, Demar M, Carme B, Darde ML. Genetic diversity, clonality and sexuality in Toxoplasma gondii. Int J Parasitol. 2004;34:1185–1196. - PubMed
-
- Bohne W, Gross U, Ferguson DJ, Heesemann J. Cloning and characterization of a bradyzoite-specifically expressed gene (hsp30/bag1) of Toxoplasma gondii, related to genes encoding small heat-shock proteins of plants. Mol Microbiol. 1995;16:1221–1230. - PubMed
-
- Bohne W, Roos DS. Stage-specific expression of a selectable marker in Toxoplasma gondii permits selective inhibition of either tachyzoites or bradyzoites. Mol Biochem Parasitol. 1997;88:115–126. - PubMed
-
- Bohne W, Wirsing A, Gross U. Bradyzoite-specific gene expression in Toxoplasma gondii requires minimal genomic elements. Mol Biochem Parasitol. 1997;85:89–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
