

PAP- and GLD-2-type poly(A) polymerases are required sequentially in cytoplasmic polyadenylation and oogenesis in Drosophila
- PMID: 18434412
- PMCID: PMC9154023
- DOI: 10.1242/dev.021444
PAP- and GLD-2-type poly(A) polymerases are required sequentially in cytoplasmic polyadenylation and oogenesis in Drosophila
Abstract
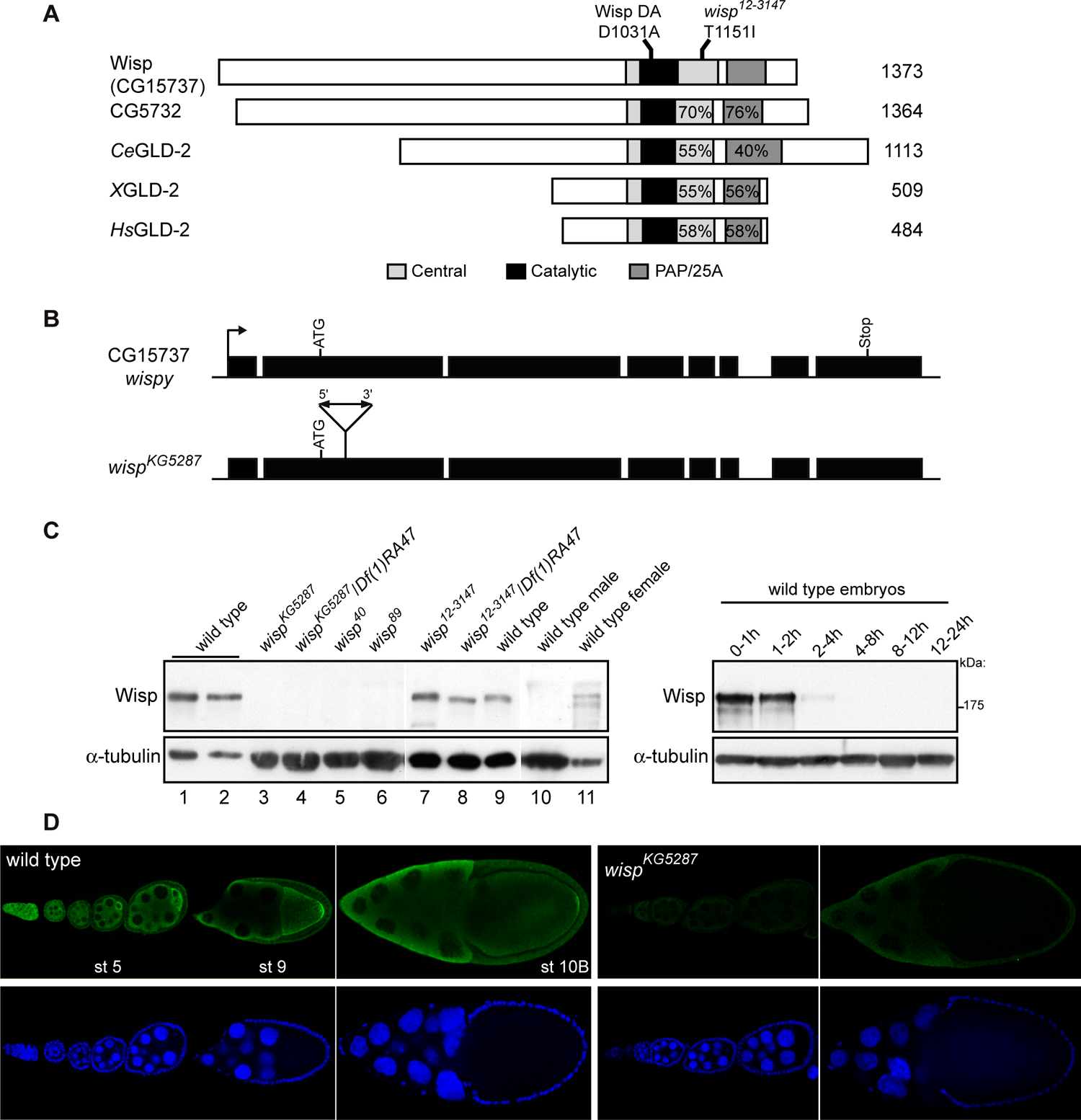
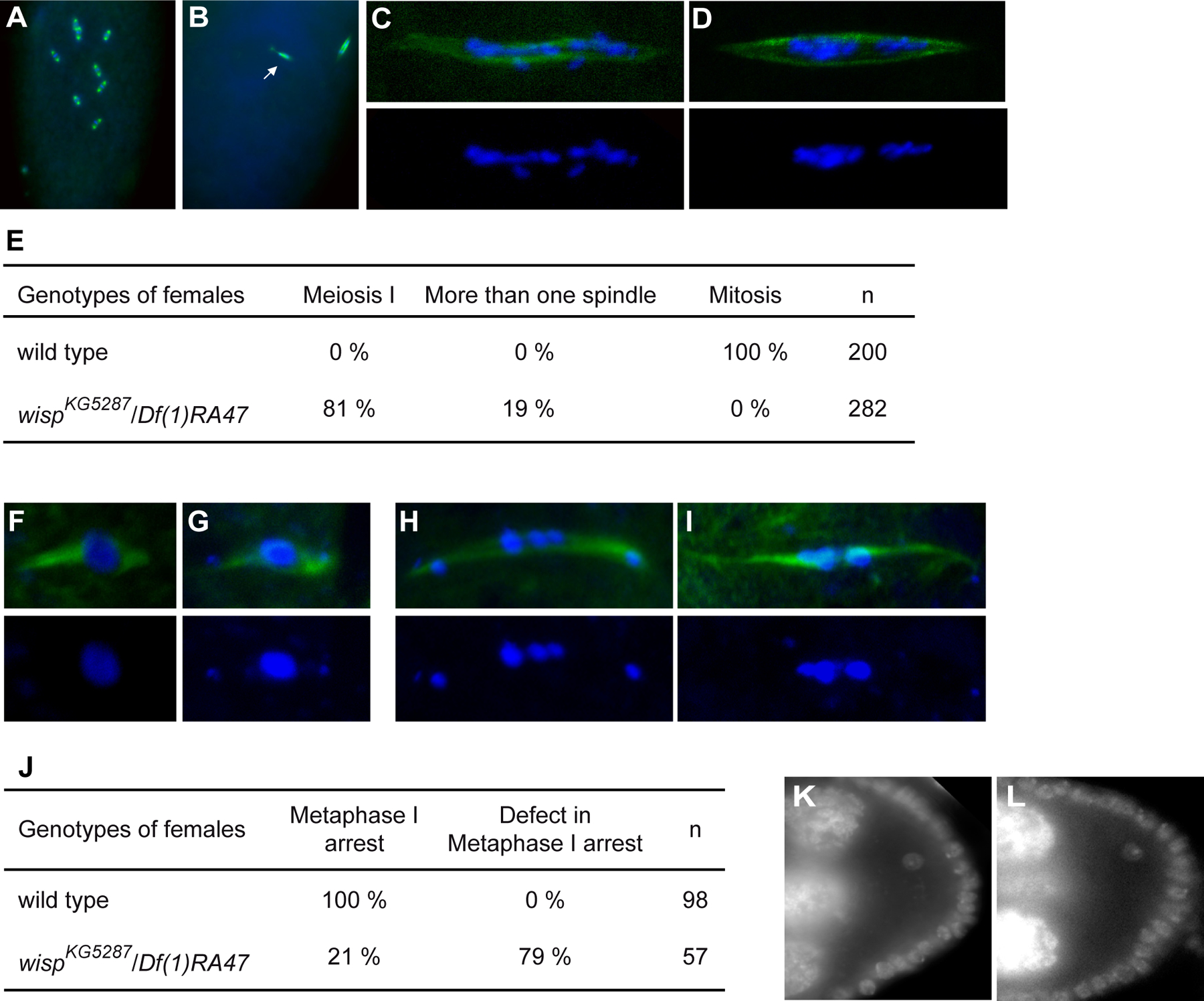
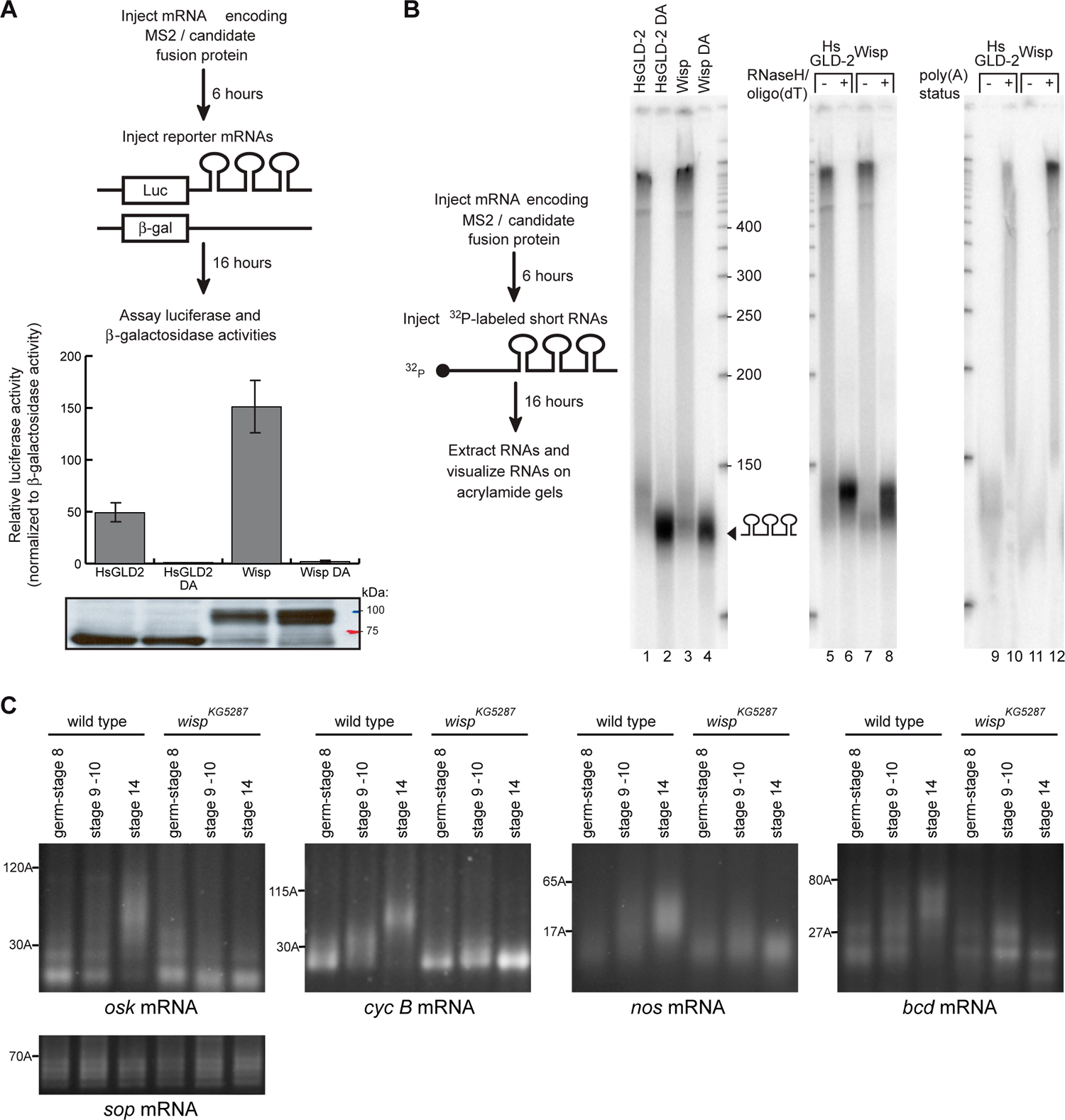
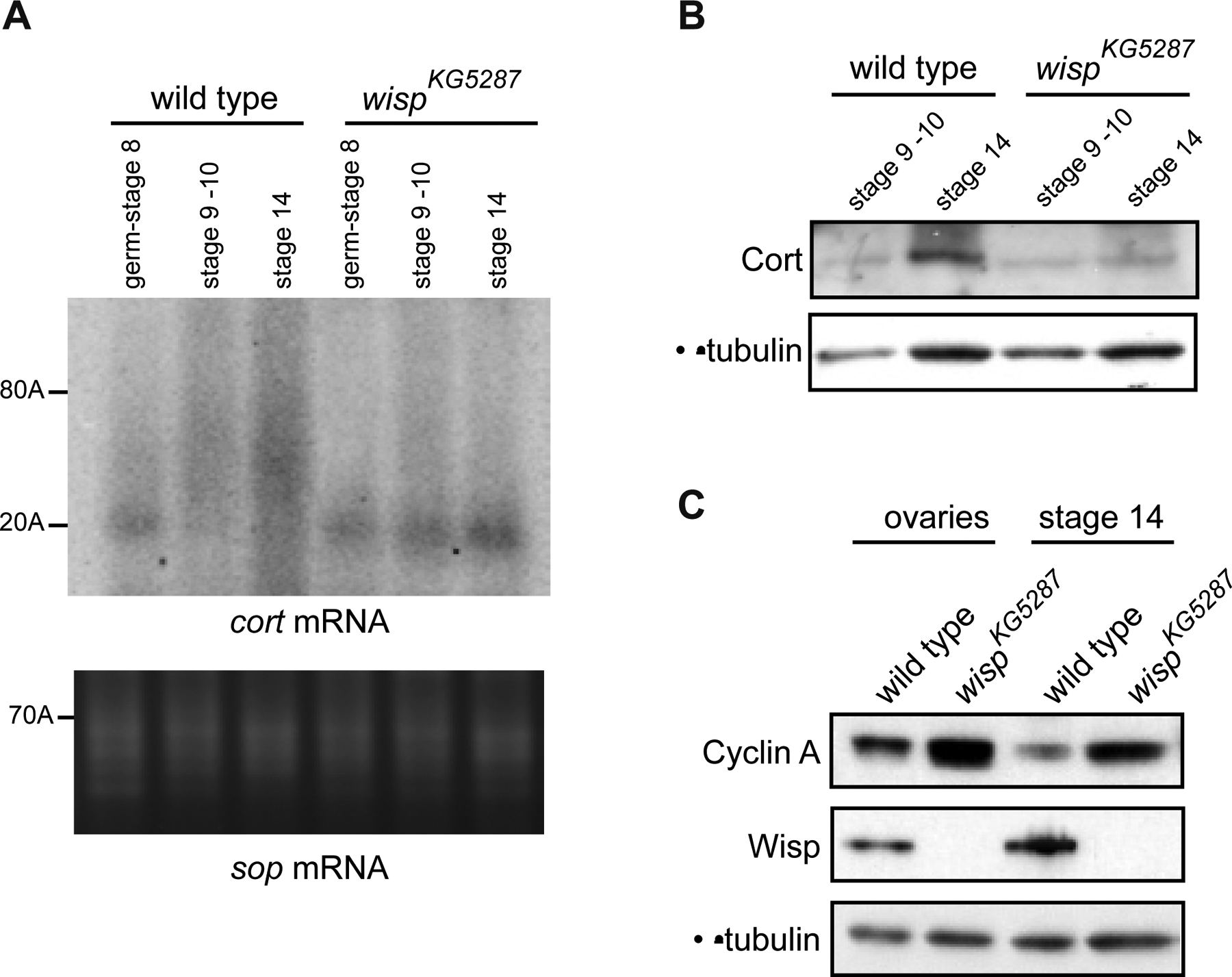
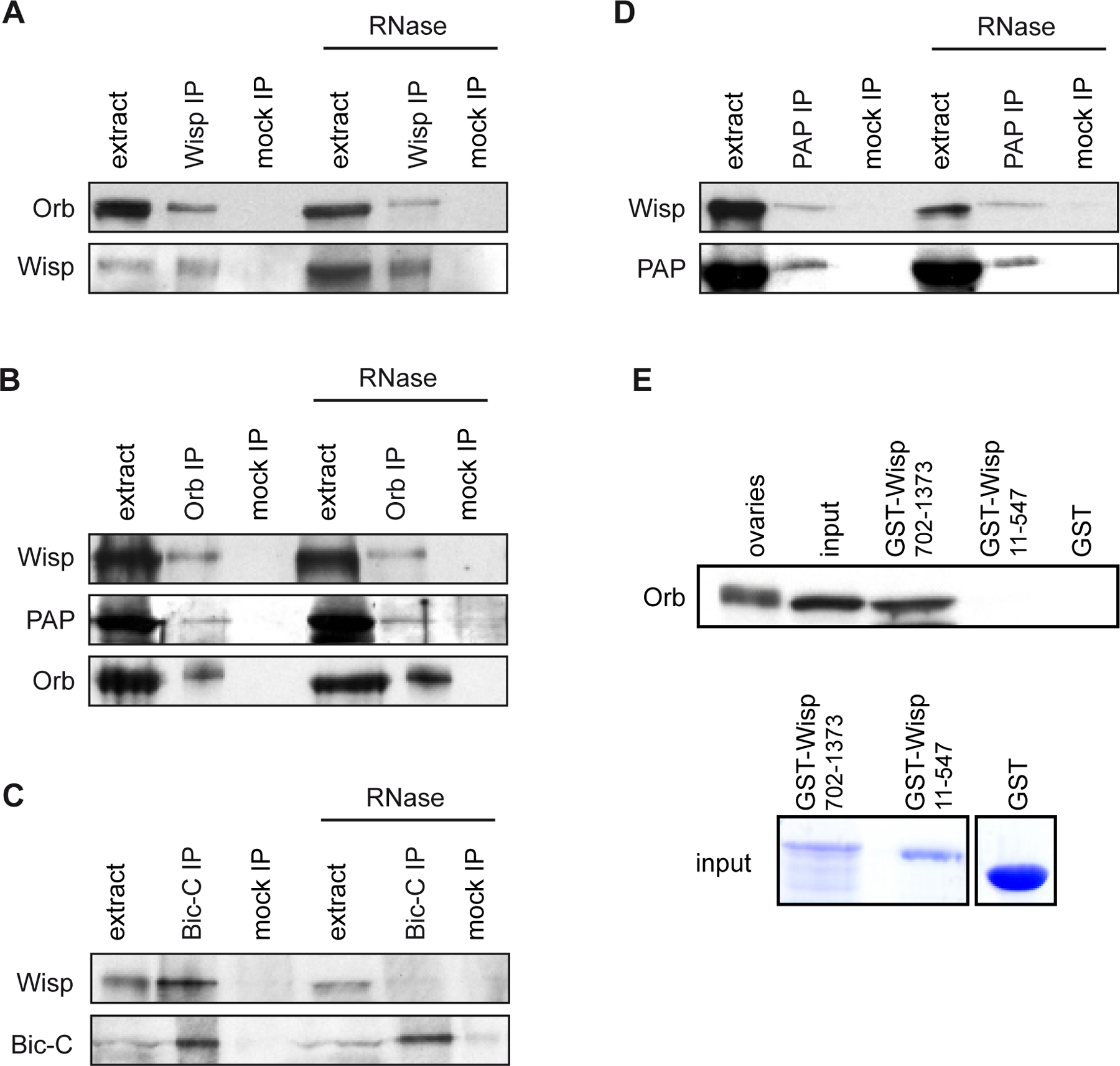
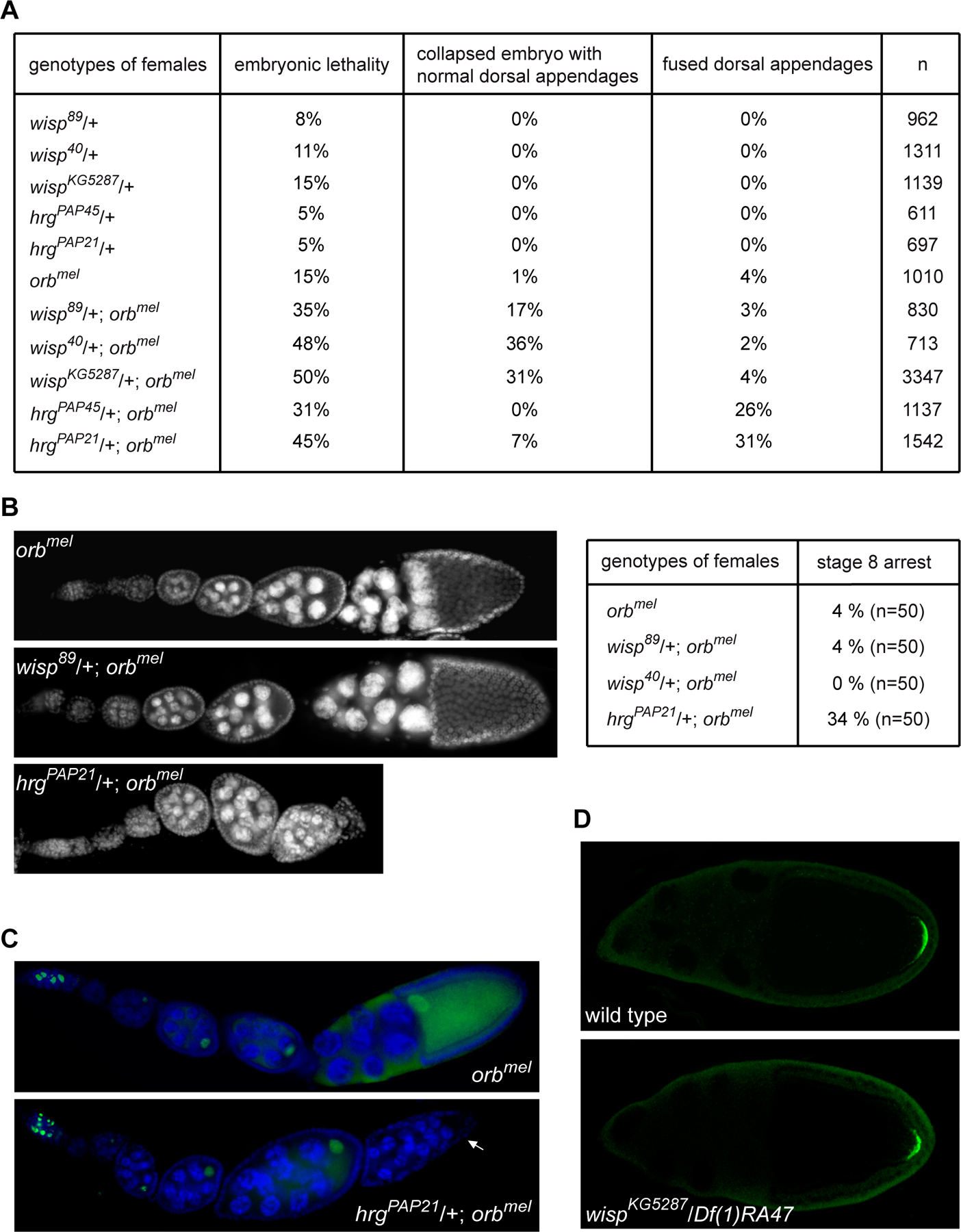
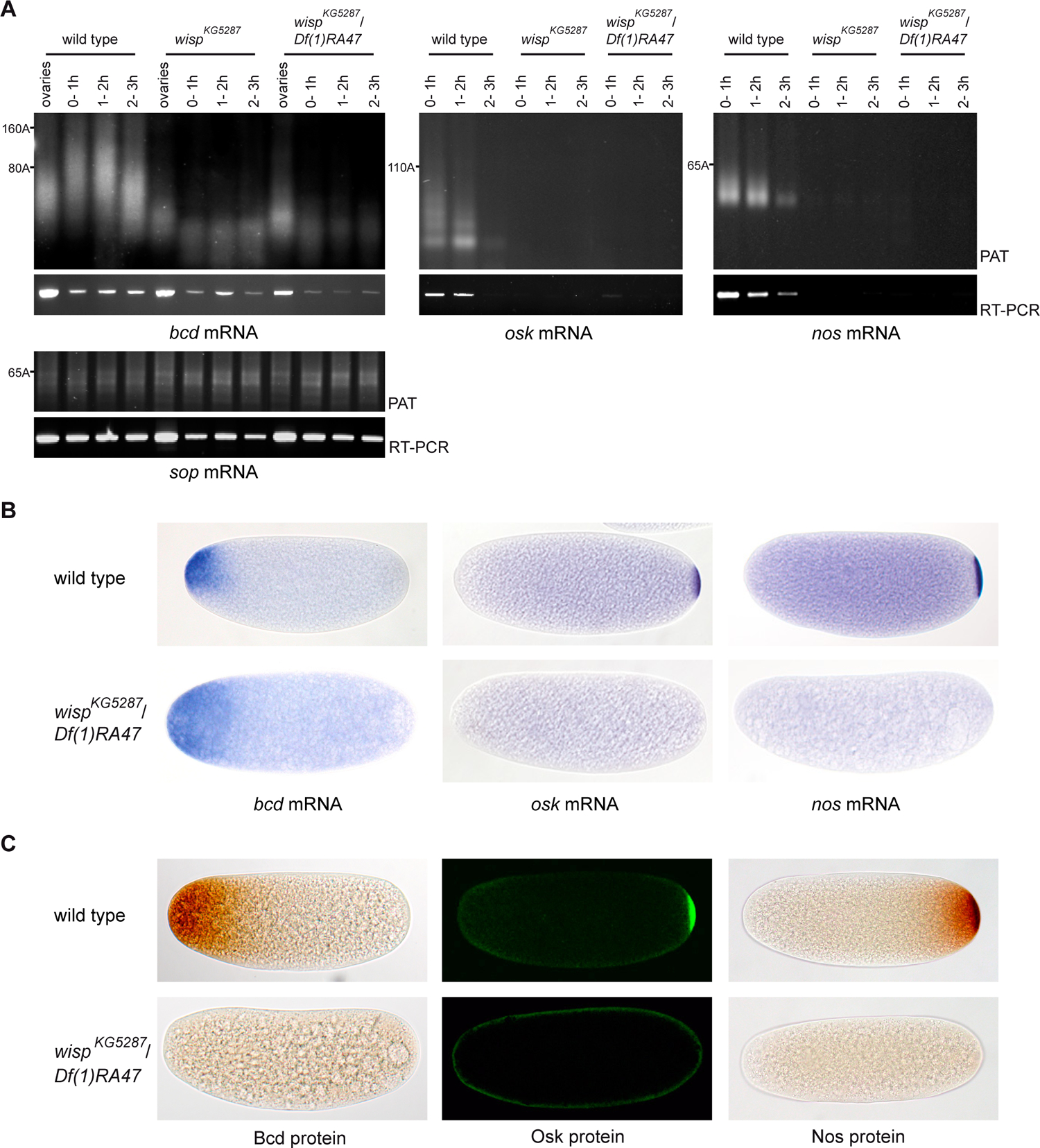
Cytoplasmic polyadenylation has an essential role in activating maternal mRNA translation during early development. In vertebrates, the reaction requires CPEB, an RNA-binding protein and the poly(A) polymerase GLD-2. GLD-2-type poly(A) polymerases form a family clearly distinguishable from canonical poly(A) polymerases (PAPs). In Drosophila, canonical PAP is involved in cytoplasmic polyadenylation with Orb, the Drosophila CPEB, during mid-oogenesis. We show that the female germline GLD-2 is encoded by wispy. Wispy acts as a poly(A) polymerase in a tethering assay and in vivo for cytoplasmic polyadenylation of specific mRNA targets during late oogenesis and early embryogenesis. wispy function is required at the final stage of oogenesis for metaphase of meiosis I arrest and for progression beyond this stage. By contrast, canonical PAP acts with Orb for the earliest steps of oogenesis. Both Wispy and PAP interact with Orb genetically and physically in an ovarian complex. We conclude that two distinct poly(A) polymerases have a role in cytoplasmic polyadenylation in the female germline, each of them being specifically required for different steps of oogenesis.
Figures
References
-
- Barnard DC, Ryan K, Manley JL and Richter JD (2004). Symplekin and xGLD-2 are required for CPEB-mediated cytoplasmic polyadenylation. Cell 119, 641–51. - PubMed
-
- Benoit B, Mitou G, Chartier A, Temme C, Zaessinger S, Wahle E, Busseau I and Simonelig M (2005). An essential cytoplasmic function for the nuclear poly(A) binding protein, PABP2, in poly(A) tail length control and early development in Drosophila. Dev Cell 9, 511–522. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
