

The PIP2 binding mode of the C2 domains of rabphilin-3A
- PMID: 18434502
- PMCID: PMC2386734
- DOI: 10.1110/ps.073326608
The PIP2 binding mode of the C2 domains of rabphilin-3A
Abstract
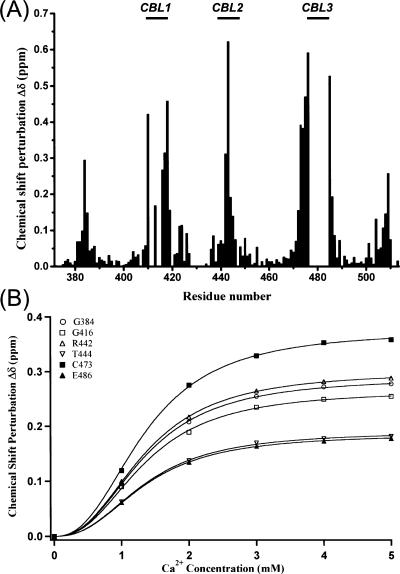
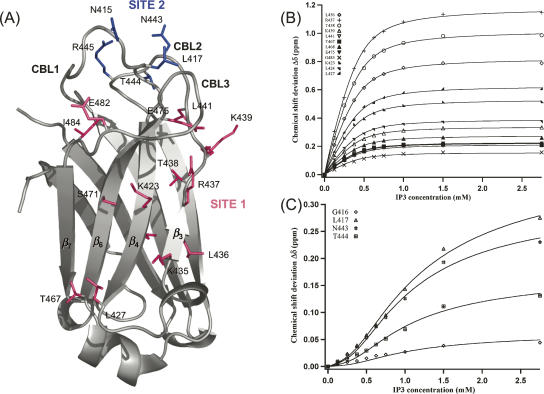
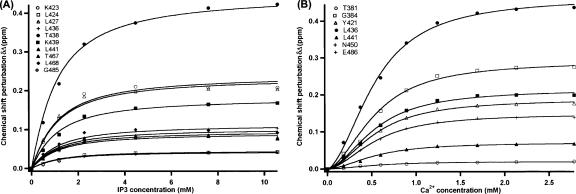
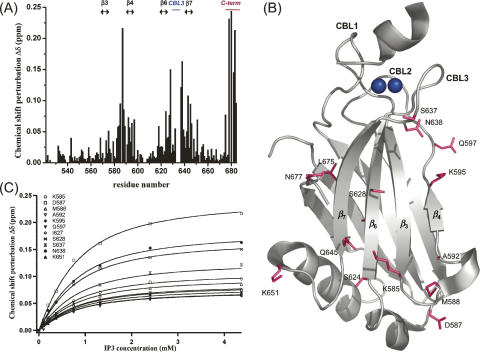
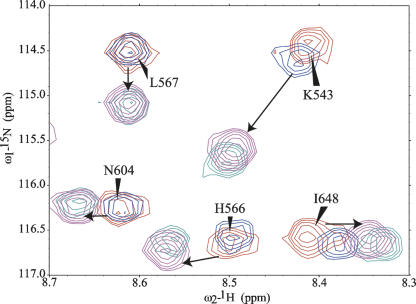
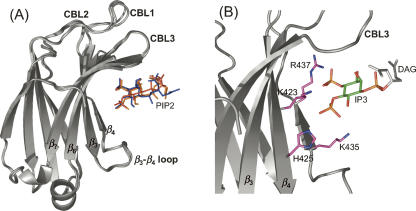
Phosphatidylinositol-4,5-bisphosphate (PIP2) is a key player in the neurotransmitter release process. Rabphilin-3A is a neuronal C2 domain tandem containing protein that is involved in this process. Both its C2 domains (C2A and C2B) are able to bind PIP2. The investigation of the interactions of the two C2 domains with the PIP2 headgroup IP3 (inositol-1,4,5-trisphosphate) by NMR showed that a well-defined binding site can be described on the concave surface of each domain. The binding modes of the two domains are different. The binding of IP3 to the C2A domain is strongly enhanced by Ca(2+) and is characterized by a K(D) of 55 microM in the presence of a saturating concentration of Ca(2+) (5 mM). Reciprocally, the binding of IP3 increases the apparent Ca(2+)-binding affinity of the C2A domain in agreement with a Target-Activated Messenger Affinity (TAMA) mechanism. The C2B domain binds IP3 in a Ca(2+)-independent fashion with low affinity. These different PIP2 headgroup recognition modes suggest that PIP2 is a target of the C2A domain of rabphilin-3A while this phospholipid is an effector of the C2B domain.
Figures

References
-
- Auguin, D., Barthe, P., Royer, C., Stern, M.H., Noguchi, M., Arold, S.T., Roumestand, C. Structural basis for the co-activation of protein kinase B by T-cell leukemia-1 (TCL1) family proto-oncoproteins. J. Biol. Chem. 2004;279:35890–35902. - PubMed
-
- Bai, J., Tucker, W.C., Chapman, E.R. PIP2 increases the speed of response of synaptotagmin and steers its membrane-penetration activity toward the plasma membrane. Nat. Struct. Mol. Biol. 2004;11:36–44. - PubMed
-
- Baldini, G., Martelli, A.M., Tabellini, G., Horn, C., Machaca, K., Narducci, P., Baldini, G. Rabphilin localizes with the cell actin cytoskeleton and stimulates association of granules with F-actin cross-linked by α-actinin. J. Biol. Chem. 2005;280:34974–34984. - PubMed
-
- Biadene, M., Montaville, P., Sheldrick, G.M., Becker, S. Structure of the C2A domain of rabphilin-3A. Acta Crystallogr. D Biol. Crystallogr. 2006;62:793–799. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
