

Multiple modes of network homeostasis in visual cortical layer 2/3
- PMID: 18434516
- PMCID: PMC2655203
- DOI: 10.1523/JNEUROSCI.5298-07.2008
Multiple modes of network homeostasis in visual cortical layer 2/3
Abstract
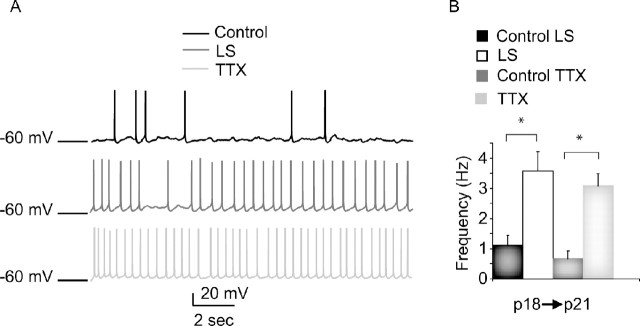
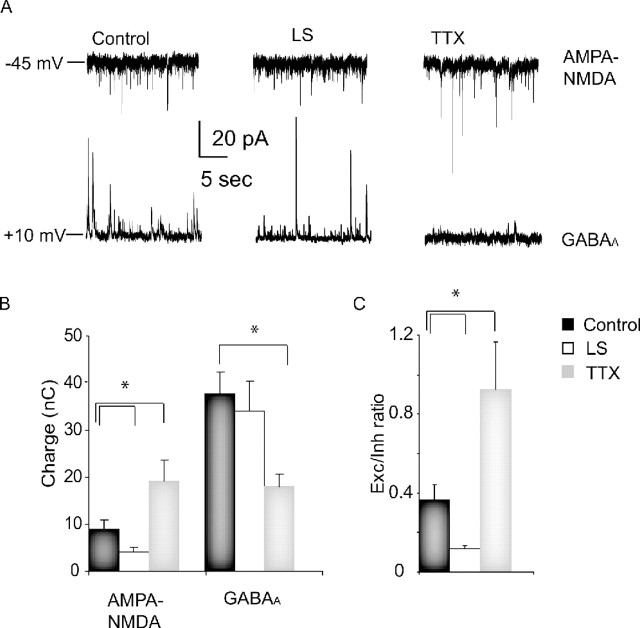
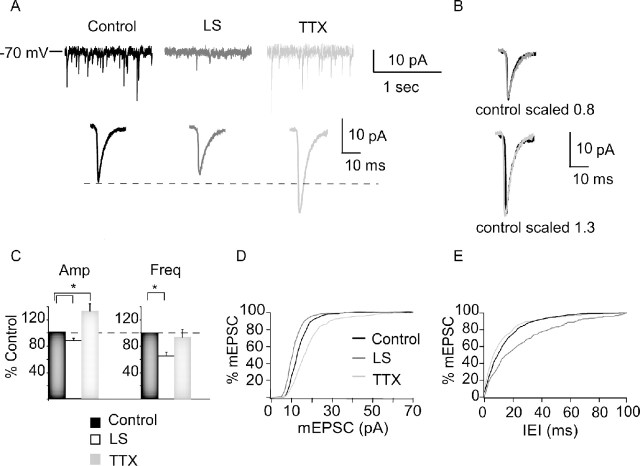
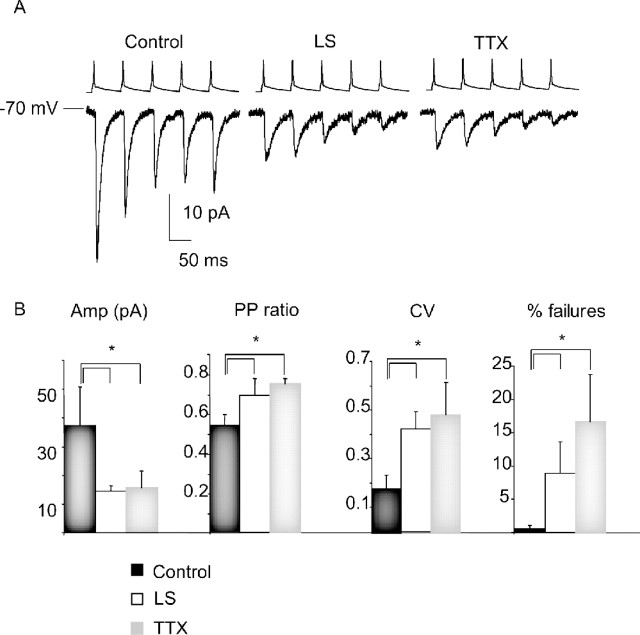
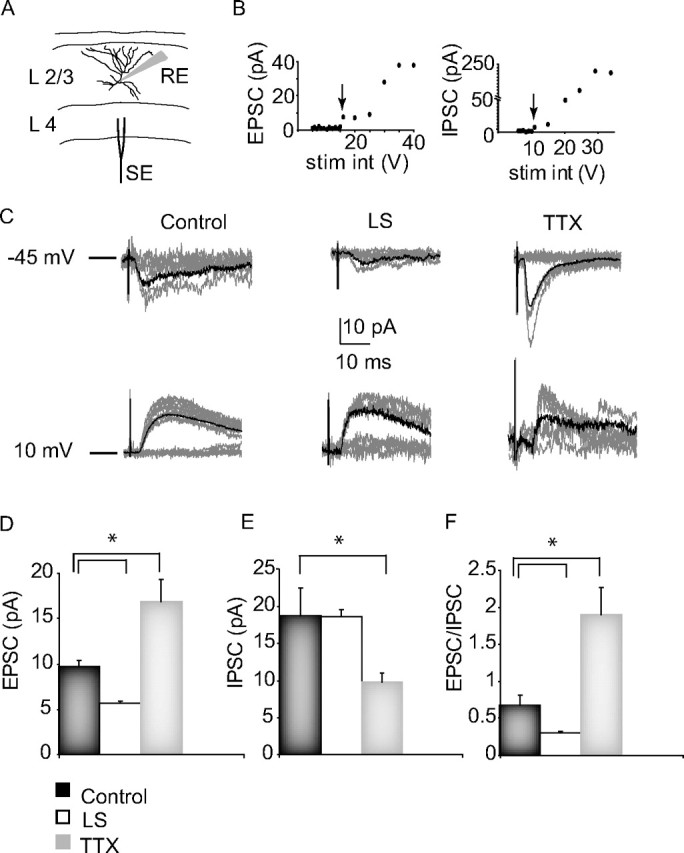
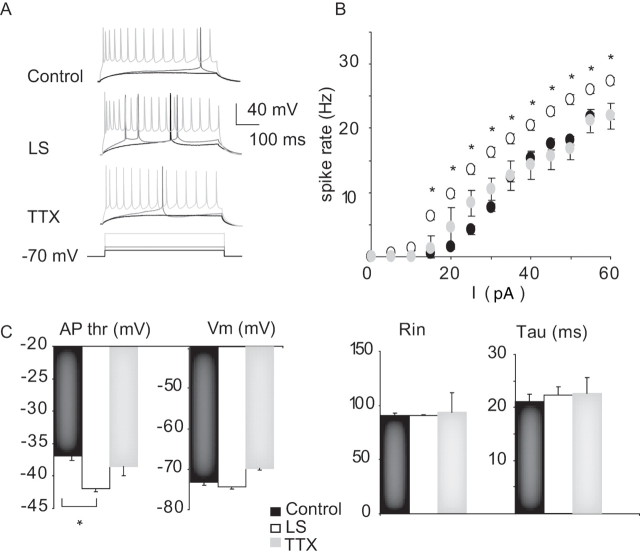
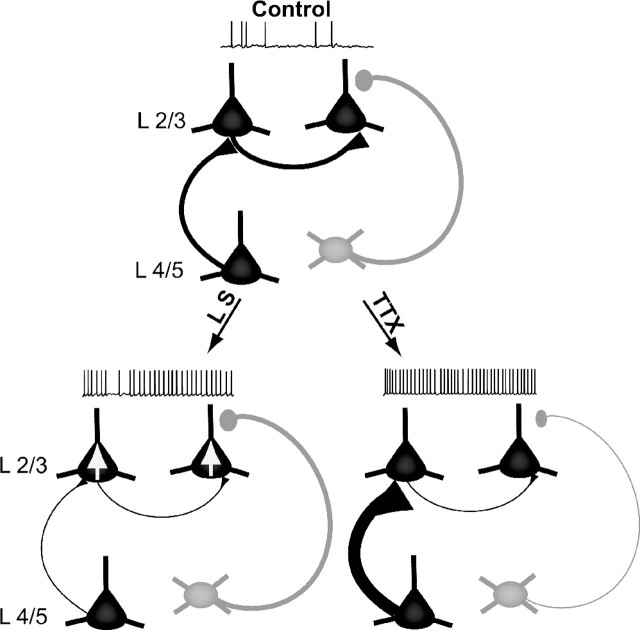
Sensory experience is crucial for shaping the cortical microcircuit during development and is thought to modify network function through several forms of Hebbian and homeostatic plasticity. Where and when these different forms of plasticity are expressed at particular synapse types within cortical microcircuits, and how they interact, is poorly understood. Here we investigated how two different visual deprivation paradigms, lid suture (LS) and intraocular TTX, affect the local microcircuit within layer 2/3 of rat visual cortex during the classical critical period for visual system plasticity. Both forms of visual deprivation produced a compensatory increase in the spontaneous firing of layer 2/3 pyramidal neurons in acute slices derived from monocular visual cortex. TTX increased spontaneous activity through an increase in the excitation/inhibition (E/I) balance within layer 2/3. In contrast, LS decreased the E/I balance by strongly depressing excitatory transmission, and the homeostatic increase in spontaneous activity was instead achieved through an increase in the intrinsic excitability of layer 2/3 pyramidal neurons. The microcircuit in layer 2/3 can thus use different forms of homeostatic plasticity to compensate for the loss of visual drive, depending on the specific demands produced by visual experience. The existence of multiple, partially redundant forms of homeostatic plasticity may ensure that network compensation can be achieved in response to a wide range of sensory perturbations.
Figures
References
-
- Aizenman C, Akerman C, Jensen K, Cline H. Visually driven regulation of intrinsic neuronal excitability improves stimulus detection in vivo. Neuron. 2003;39:831–842. - PubMed
-
- Allen CB, Celikel T, Feldman DE. Long-term depression induced by sensory deprivation during cortical map plasticity in vivo. Nat Neurosci. 2003;6:291–299. - PubMed
-
- Burkhalter A. Intrinsic connections of rat primary visual cortex: laminar organization of axonal projections. J Comp Neurol. 1999;279:171–186. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources